
חזית המחקר
השפעת נבירה של חזירי הבר בקרקע על חברת הצמחים העשבוניים
אבי פרבולוצקי המכון למדעי הצמח, המחלקה לגידולי שדה ומשאבי טבע, ִמנהל המחקר החקלאי ([email protected])
גידי נאמן החוג להוראת המדעים – המחלקה לביולוגיה, אוניברסיטת חיפה-אורנים

חזיר בר ברמת הנדיב. צולם באמצעות מצלמה סטטית המוצבת בשטח ומזהה תנועה על צירי תנועה אוכלוסיית חזירי הבר | צילום: ביל וודלי
מאת
גיא דוברת
החוג לביולוגיה אבולוציונית וסביבתית, אוניברסיטת חיפהאבי פרבולוצקי
המכון למדעי הצמח, המחלקה לגידולי שדה ומשאבי טבע, ִמנהל המחקר החקלאיגידי נאמן
החוג להוראת המדעים – המחלקה לביולוגיה, אוניברסיטת חיפה-אורניםמאת
גיא דוברת
החוג לביולוגיה אבולוציונית וסביבתית, אוניברסיטת חיפהאבי פרבולוצקי
המכון למדעי הצמח, המחלקה לגידולי שדה ומשאבי טבע, ִמנהל המחקר החקלאיגידי נאמן
החוג להוראת המדעים – המחלקה לביולוגיה, אוניברסיטת חיפה-אורניםחזיר הבר הוא היונק היבשתי הטבעי הגדול ביותר ששרד בישראל. פעילות חזירי הבר ניכרת בשטחים טבעיים וחקלאיים ובתחום יישובי אדם. נבירה של חזירי בר בקרקע גורמת להפרת האיזון של המערכת האקולוגית ואמורה להביא לשינוי משמעותי במאפייני החברה העשבונית וביסודות אקולוגיים נוספים. במחקר זה בחנו את השפעת הנבירה של חזירי הבר על חברת הצמחים העשבוניים בכתמים פתוחים בגריגה ים תיכונית ברמת הנדיב שבדרום הכרמל. בחנו ארבעה טיפולים שייצגו עוצמות ומועדי נבירה שונים: נבירת קיץ, נבירת חורף, שטח מגודר ללא נבירה ושטח פתוח ללא נבירה. הנבירה מתקיימת במשך כל השנה. בחורף ובאביב היה היקף הנבירה כ-4% מכלל השטח הניתן לנבירה, ובקיץ כ-2%, כתלות בבית הגידול. מצאנו כי הנבירה גרמה לשינוי מוגבל במבנה של חברת הצמחים העשבוניים ובהרכבה בהשוואה לשטחים מגודרים שבהם נמנעה הנבירה. נבירת קיץ, טרם נביטה, העלתה את מגוון המינים אך לא שינתה את היבול הצמחי. נבירת חורף, בתקופת הצמיחה, הפחיתה את היבול הצמחי אך לא שינתה את עושר המינים ואת צפיפות הפרטים. נבירות בעוצמות שונות, בהיקפים שונים ובעונות שונות יוצרות כתמים בעלי תנאי קרקע ותחרות שונים, אך אין בהם כדי לגרום לכתמיות ברורה וקבועה בנוף הצומח העשבוני. הניגוד בין היקף הנבירה הגדול וביטויו הנופי לבין ההשפעה האקולוגית הכמעט השולית על חברת הצומח העשבוני יכול להיות מוסבר בכך שבבתי גידול טבעיים כמו רמת הנדיב, מותאמת החברה העשבונית להפרה על-ידי נבירה.
על קצה המזלג
השפעתם של חזירי הבר על שטחים חקלאיים ואף בתוך יישובים כפריים מוכרת כמזיקה. חזירי בר נוהגים לחפור ולנבור בקרקע במטרה לחפש מזון. פעילות נבירה זו מתקיימת גם בשטחים הטבעיים שחיים בהם חזירי הבר. כיונק היבשתי הטבעי הגדול בישראל, יש לחזירי הבר היכולת להפר את האיזון של המערכת האקולוגית ואף לשנותה. במחקר שנערך בפארק רמת הנדיב שבדרום הכרמל, נבחנה השפעת הנבירה בקרקע על הצמחייה העשבונית הים תיכונית (צמחים נמוכים שמחזור החיים שלהם קצר, ואינו עולה על שנה). נמצא שבעוד שהנבירה בולטת מאוד בנוף ואף משתרעת על פני שטחים נרחבים, הרי שהשפעתה האקולוגית על הצמחייה העשבונית הייתה קטנה בלבד, ואף נמצאה בה תועלת מבחינת שימור מגוון מיני הצומח העשבוני. ייתכן שהפער הזה נוצר מכיוון שמערכות טבעית כגון זו שבאזור הנבדק הספיקו לעבור תהליכי הסתגלות אִטיים (מבחינת הרכב המינים ויכולתם להתאושש) להפרעות שיוצרות נבירות חזירי הבר בקרקע, בדומה להשפעות של רעיית צאן ובקר. מסיבות אלה, חזירי הבר אינם יכולים להיחשב 'מהנדסי סביבה' באזור הים תיכוני. הנזקים שגורמים חזירי בר בעקבות חדירה לאזורים חקלאיים ומיושבים יוצרת לחץ לקבלת החלטות שיביאו לצמצום השפעתם, אך חשוב שתילקח בחשבון גם תרומתם בפועל לשימור מגוון המינים של מיני עשב. יש חשיבות לניטור של אוכלוסיות חזירי הבר ושל השפעתן על מערכות אקולוגיות – בשטחים פתוחים ובמסדרונות האקולוגיים שביניהם – כדי ליצור בסיס לקבלת החלטות מושכלת
מערכת אקולוגיה וסביבה
הלכה למעשה
מבוא
חזיר הבר (.Sus scrofa L) הוא יונק גדול בעל תפוצה גאוגרפית רחבה, המופיע במגוון גדול של בתי גידול [12, 25]. תפוצתו המקורית היא אירואסיינית, אולם בעקבות פעולות האדם הוא נפוץ כיום ברוב העולם. בישראל נמצא חזיר הבר מתת-המין S. scrofa lybicus, הנפוץ מהים השחור בצפון, ועד ישראל שהיא גבול תפוצתו הדרומי.
חזירי הבר נוברים בקרקע לשיחור מזון המורכב בעיקר מחלקי צמחים [7], מפטריות ומחסרי חוליות [12, 25]. הנבירה מבוצעת על-ידי נעיצת הלסת התחתונה בקרקע והרמתה תוך הפיכת כמות גדולה של קרקע. הנבירה חושפת את הקרקע ופוגעת בצמחייה, והיקפה משתנה בין שנים ועונות [28], ובין חברות צמחים וסוגי קרקע [6, 11]. חזירי בר פעילים גם בסמוך למגורי אדם וגורמים נזקים קשים לחקלאות [6, 12]. מחקרים רבים שבחנו את השפעתה של נבירה המבוצעת על-ידי חזירים בוצעו במקומות שבהם הם מין גר ופליט תרבות [9, 11, 20, 23, 27] ורק מעטים נערכו בבית גידולם הטבעי [14].
אמנם אין נתונים כמותיים על אוכלוסיות חזירי הבר בישראל, אך נראה כי בעשורים האחרונים הייתה עלייה חדה במספרם. הסיבות לכך הן התפשטות שטחי חקלאות השלחין והתמעטותם של טורפים טבעיים. חזירים מלווים את מרבית שטחי חקלאות בישראל ולכן הוכרזו כבעל חיים מזיק המותר לציד.
שטחים טבעיים במערכת האקולוגית הים תיכונית מורכבים מפסיפס של כתמי צומח בגדלים שונים ובתדירות משתנה. כתמים אלו הם תוצאה של הבדלים בפיזור המרחבי והעתי של משאבים כמו מים, אור ומינרלים, הנגרמים בין היתר גם על-ידי הפרות שונות. כתוצאה מכך חברות הצומח הן בעלות מבנה הטרוגני [18, 22]. בעלי חיים שונים יוצרים הפרעות בקנה מידה קטן ומשפיעים בכך ישירות על המבנה ועל הפיזור המרחבי של הכתמים בנוף, ובאופן עקיף גם על הרכב המינים ועל מגוונם [3, 26, 29]. נמצא שגם צמחים מעוצים משפיעים על פיזור המשאבים במרחב ובעקיפין על ההרכב של חברות אקולוגיות שונות [2, 24]. אורגניזם (יצור) המשפיע בצורה ניכרת מעצם נוכחותו או פעילותו, שלא דרך מארג המזון, על פיזור המשאבים והאורגניזמים בסביבה מוגדר כ‘מהנדס סביבה‘ [15, 16, 24].
פעילות חזירי הבר ניכרת בשטחים חקלאיים וטבעיים בעיקר בחורף, בעונת הצמיחה של הצמחים העשבוניים. כתוצאה מכך עלה חשש שפעילות חזירי הבר עלולה לפגוע במגוון המינים העשבוניים שהם מרכיב חשוב במגוון הצמחים בישראל. מטרות המחקר הן לבדוק את השפעתה של הנבירה שמבצעים חזירי הבר על המגוון של הצמחייה העשבונית ועל הרכבה בכתמים הפתוחים בחורש, ולבדוק את היקפי הנבירה לאורך השנה באזור הים תיכוני.
שיטות
המחקר נערך בפארק רמת הנדיב (נ.צ. 7181/1945) ברום של 120 מטר מעל לפני הים. הקרקע היא טרה רוסה על גבי תצורות סלע גירניות [1]. האקלים ים תיכוני עם 550 מ“מ גשם בשנה בממוצע. פארק רמת הנדיב מכוסה ברובו גריגה ים תיכונית (שיחים בגובה של עד כמטר וחצי) ואזורים הנטועים בחורשות מחטניים.
השפעתן של נבירות שמבצעים חזירי הבר בקרקע
כדי לייצג מגוון בתי גידול בחרנו שישה אתרים במפנה צפוני, שישה במפנה דרומי ושמונה על הרמה. במהלך 2007–2008 סומנו בכל אתר נבחר ארבע חלקות טיפול (בגודל 90×90 ס“מ). החלקות נמצאו באזורים שהיו בהם נבירות טבעיות בשטח, וכן נבחרו חלקות שנמנעה בהן נבירה. מיקום ההחלקות נקבע כך שיהיו סמוכות זו לזו ובהן הטיפולים הבאים:
- בקרה בעזרת גידור: חלקה ללא נבירה, שגידרנו ביולי 2007 למניעת פעילות של חזירי בר.
- נבירות קיץ: חלקה שחזירי בר נברו בה בין מאי לספטמבר 2007 ואז גודרה למניעת נבירות חוזרות.
- נבירות חורף: בפברואר 2008 איתרנו חלקות שהיו בהן נבירות טריות שמופו וסומנו אך לא גודרו.
- בקרה ללא גידור: בסמוך לחלקות הבקרה המגודרות שסימנו ביולי 2007, איתרנו חלקות שהיו חשופות לנבירה שסומנו ולא גודרו. בחלקות אלה ביצענו מעקב דו-שבועי אחר נבירות. חלקות שלא נברו בהן הושוו לחלקות הבקרה המגודרות כדי לקבוע את השפעת הגידור עצמו על הצומח.
בדקנו את השפעת הנבירה על עושר מיני הצומח, על מגוונם ועל הביומסה העשבונית, בכתמים פתוחים בין שיחי הגריגה המכוסים באביב בצמחייה עשבונית. את כל החלקות דגמנו בשיא התפתחות הצומח העשבוני (30.3.2008–2.4.2008).
לדגימת הצמחים העשבוניים בכל חלקת ניסוי (גדורה או מסומנת), השתמשנו בריבוע בגודל 20×20 ס“מ שהונח בנקודה מייצגת בכל טיפול. קצרנו את כל הצמחים העשבוניים בריבוע ולקחנו אותם למעבדה לשם הגדרה, ספירת הפרטים מכל מין, ייבוש ושקילה לקביעת הביומסה.
שינוים בהיקף הנבירה ובעוצמתה
כדי להעריך את השינויים העונתיים בהיקף הנבירה ביצענו שלושה סקרים: בקיץ, בחורף ובאביב. הסקרים נפרסו על פני 1,000 דונם של גריגה וחורשות נטועות. בכל עונה דגמנו 73 נקודות קבועות שמיקומן המדויק נקבע מראש, באופן מבוקר, בעזרת GPS. בכל נקודה הונחו שני חתכים צולבים באורך של 20 מטר. בכל חתך נמדד אורך שטח הנבירה שנמצא מתחת לסרט המדידה ואורך שטח הקרקע הניתן לנבירה שאינו כולל סלעים וצומח מעוצה. אורך שטח הנבירה בפועל יחסית לאורך השטח הניתן לנבירה ייצג את אחוז שטח הנבירות בחתך. עוצמת הנבירה נאמדה ב-40 חלקות של טיפולי הנבירה בלבד, שם גם בוצעה הערכה ויזואלית של עומק הנבירה וחלוקה לשלוש עוצמות של נבירה.
ניתוח הנתונים
ניתוח הנתונים בוצע בתכנת SPSS – SPSS 14 for Windows SPSS Inc. 2005. כל הנתונים (שפע, עושר ומגוון מינים וביומסה) שנאספו בחלקות ובאתרים השונים הושוו בעזרת מבחן שונויות למדידות חוזרות (Repeated measures ANOVA) ומבחן Post-hock Bonferroni. נוסף על כך הושוו תוצאות הבדיקות בחלקות הבקרה המגודרות לאלו של חלקות הבקרה הבלתי מגודרות בעזרת מבחן T מזווג (Paired-samples t-test). לחישוב מגוון המינים השתמשנו במדד המִגוון של שאנון Shannon) 1948). לבדיקת הקשר בין הרכב מיני הצמחים לטיפולים השונים השתמשנו בתכנת האורדינציה [4, 5] CANOCO 4.5 המבצעת מבחן Monte-Carlo לנתונים בהתייחס למינים ולמשתנים סביבתיים שנדגמו ונמדדו באותן דגימות. לבדיקת השפעתה של הנבירה על המינים הדומיננטיים ועל היחסים הכמותיים ביניהם, חושב השפע היחסי של מספר הפרטים של חמשת המינים הדומיננטיים בכל דגימה, ונערכה השוואה בין הטיפולים לגביהם. נתוני סקר היקפי הנבירה בעונות השונות ומאפייני צומח הושוו בעזרת מבחן פרידמן (Friedman test).
תוצאות
השפעת הנבירה על צפיפות הצמחים, על עושר המינים ומגוונם ועל הביומסה של הצמחים העשבוניים
במהלך דגימת הצומח אספנו 9,998 פרטים שהשתייכו ל-110 מיני צמחים עשבוניים (מתוך כ-570 מינים שזוהו ברמת הנדיב).
צפיפות הצמחים הממוצעת בחלקות הבקרה המגודרות (463 ± 3,620 פרטים למ“ר) הייתה גבוהה יותר מאשר בחלקות נבירת הקיץ (238 ± 2,188 פרטים למ“ר) ובחלקות נבירת החורף (320 ± 2,475 פרטים למ“ר). בניתוח שונויות למדידות חוזרות נמצא הבדל מובהק בין הטיפולים (RM ANOVA, F1.7,31.6=5.786, P=0.01), אולם במבחן הסטטיסטי Post-hock Bonferroni נמצא הבדל מובהק רק בין הטיפול בחלקות הבקרה המגודרות לבין נבירת הקיץ, ואילו נבירת החורף לא הייתה שונה בהשפעתה על החברה העשבונית משאר הטיפולים.
עושר המינים הממוצע בחלקות של נבירת קיץ (0.84 ± 9.65 מינים) היה גבוה מזה של חלקות נבירת החורף (0.76 ± 7.35 מינים) ומאלו של חלקות הבקרה המגודרות (0.83 ± 8.35 מינים). נמצאה השפעה מובהקת של הטיפולים על עושר המינים (RM ANOVA, F2,38=3.492, P=0.041). בבדיקה באמצעות מבחן Post-hock Bonferroni נמצא הבדל מובהק רק בין חלקות נבירת הקיץ לבין חלקות נבירת החורף, בעוד שחלקות הבקרה המגודרות לא היו שונות משאר הטיפולים.
מגוון המינים הגבוה ביותר, לפי המדד של שאנון, היה בחלקות נבירת הקיץ. בחלקות הבקרה המגודרות ובחלקות נבירת החורף היה מגוון נמוך יותר (איור 1א). נמצאה השפעה מובהקת של הטיפולים על המגוון (RM ANOVA, F2,38=7.661, P=0.01), אולם במבחן Post-hock Bonferroni נמצא הבדל מובהק רק בין חלקות נבירת הקיץ לבין חלקות נבירת החורף וחלקות הבקרה המגודרות, שלא היו שונות אלו מאלו.
לא נמצאו הבדלים בעושר המינים, במספר הפרטים ובביומסה בין חלקות הבקרה המגודרות לבין חלקות הבקרה הבלתי מגודרות.
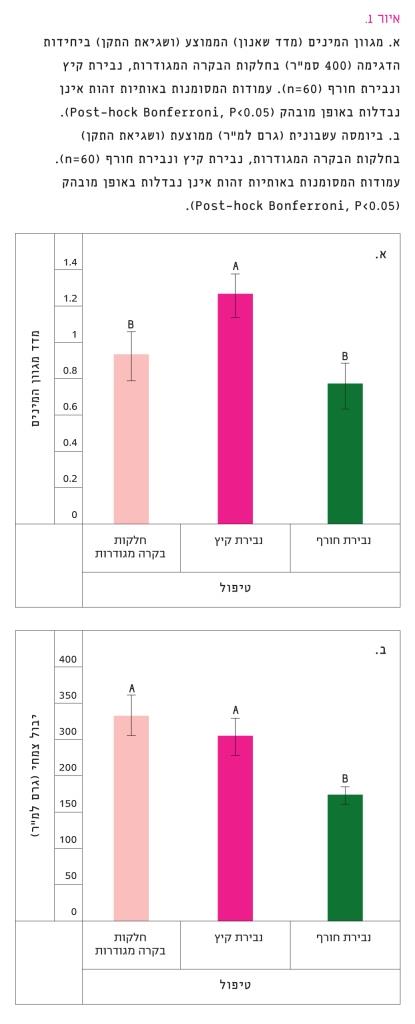
איור 1
א. מגוון המינים (מדד שאנון) הממוצע (ושגיאת התקן) ביחידות הדגימה (400 סמ"ר) בחלקות הבקרה המגודרות, נבירת קיץ ונבירת חורף
60=n. עמודות המסומנות באותיות זהות אינן נבדלות באופן מובהק (0.05>Post-hock Bonferroni, P).
ב. ביומסה עשבונית (גרם למ"ר) ממוצעת (ושגיאת התקן) בחלקות הבקרה המגודרות, נבירת קיץ ונבירת חורף
60=n. עמודות המסומנות באותיות זהות אינן נבדלות באופן מובהק (Post-hock Bonferroni, P<0.05).
מידת הדומיננטיות של המינים הנפוצים נבדקה על-ידי השוואת מספר הפרטים הממוצע של כל אחד מחמשת המינים הדומיננטיים, ללא התייחסות לזהות המינים, בכל אחד מהטיפולים (איור 2). נמצא הבדל מובהק (RM ANOVA, F1.7,31.7=6.27, P=0.007) בניתוח שונויות למדידות חוזרות, שבוצע לגבי המין הנפוץ ביותר בשלושת הטיפולים.
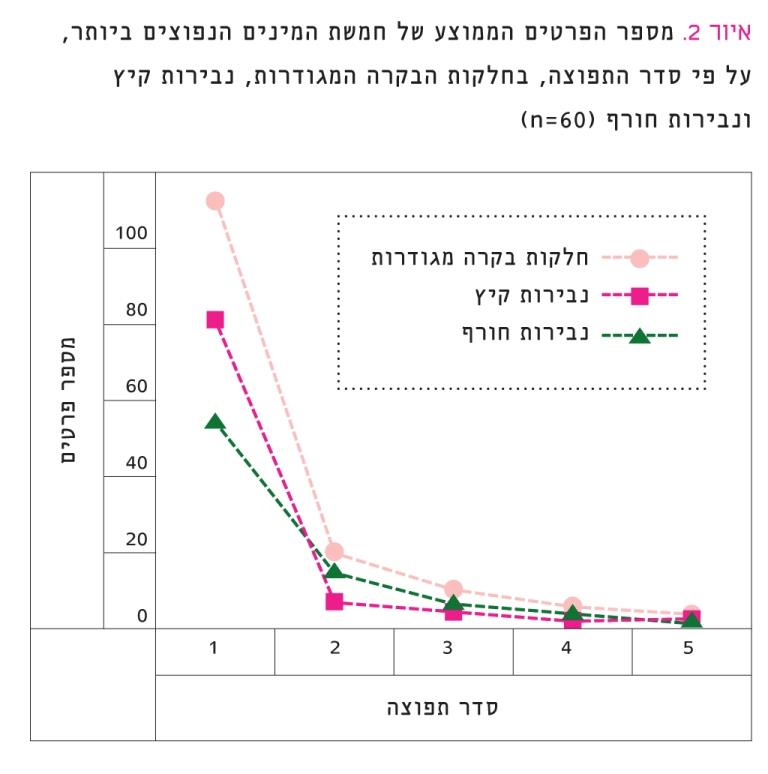
איור 2
מספר הפרטים הממוצע של חמשת המינים הנפוצים ביותר, על פי סדר התפוצה, בחלקות הבקרה המגודרות, נבירות קיץ ונבירות חורף (60=n)
הביומסה העשבונית ליחידת שטח הייתה גבוהה בחלקות הבקרה המגודרות ובחלקות נבירת הקיץ, בעוד שבחלקות נבירת החורף הייתה הנמוכה ביותר (איור 1ב). נמצאה השפעה מובהקת של הטיפולים על הביומסה העשבונית (RM ANOVA, F2,38=8.547, P=0.001). במבחן Post-hock Bonferroni נמצא הבדל מובהק רק בין חלקות נבירת החורף לבין חלקות נבירת הקיץ וחלקות הבקרה המגודרות, שלא היו שונות אלו מאלו.
בניתוח האורדינציה (באמצעות CANOCO) נבדק הקשר בין השפע של מיני הצמחים העשבוניים לטיפולים ולעוצמת הנבירה. בניתוח נכללו 44 מינים שבכל החלקות נמצאו מהם לפחות עשרה פרטים, אולם בחרנו להציג רק 24 מינים שהראו תגובה ברורה לטיפולים (איור 3). 20 המינים שהראו פחות מ-3% התאמה למודל מוקמו קרוב למוצא הצירים ולכן לא הוצגו בגרף. התוצאות נמצאו מובהקות (Monte-Carlo test, F=2.549, P=0.048) ובגרף ניתן לראות חלוקה לשלוש קבוצות: (1) מינים שהופיעו בעיקר בחלקות הבקרה ללא נבירה; (2) מינים שהופיעו בעיקר בחלקות שנעשתה בהן נבירה חלשה ובינונית; (3) מינים שהופיעו בעיקר בחלקות שעברו נבירת חורף חזקה.
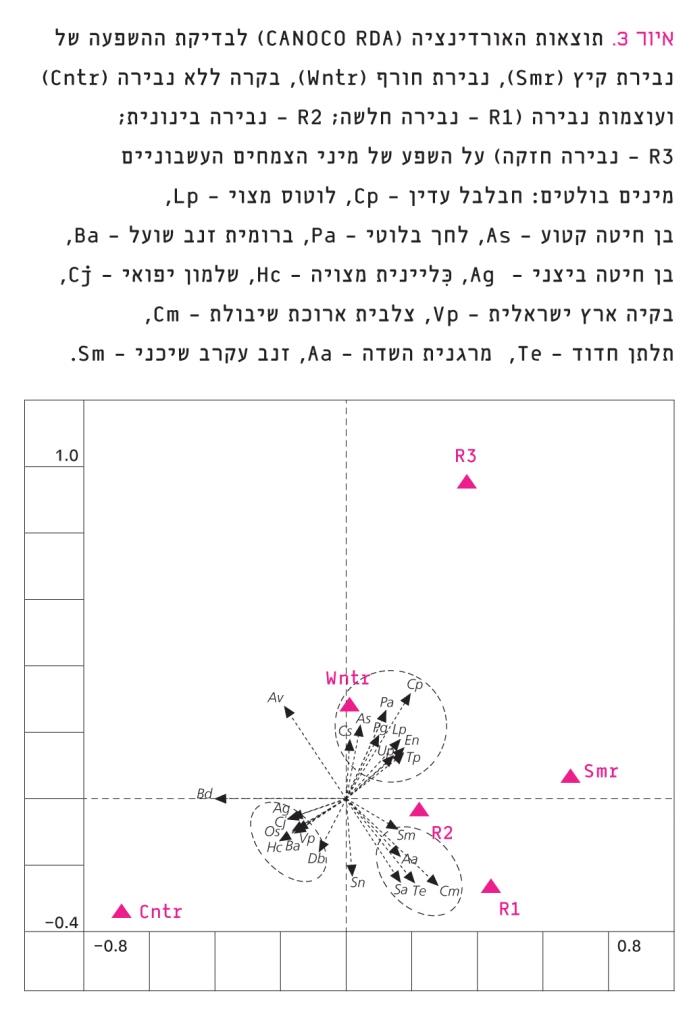
איור 3
תוצאות האורדינציה (CANOCO RDA) לבדיקת ההשפעה של נבירת קיץ (Smr), נבירת חורף (Wntr), בקרה ללא נבירה (Cntr) ועוצמות נבירה (R1 – נבירה חלשה; R2 – נבירה בינונית; R3 – נבירה חזקה) על השפע של מיני הצמחים העשבוניים
מינים בולטים: חבלבל עדין – Cp, לוטוס מצוי – Lp, בן חיטה קטוע – As, לחך בלוטי – Pa, ברומית זנב שועל – Ba, בן חיטה ביצני – Ag, ִּכליינית מצויה – Hc, שלמון יפואי – Cj, בקיה ארץ ישראלית – Vp, צלבית ארוכת שיבולת – Cm, תלתן חדוד – Te, מרגנית השדה – Aa, זנב עקרב שיכני – Sm
השנויים בהיקף הנבירה של חזירי הבר
שלושת סקרי הנבירה הראו כי שטח הנבירות בכתמים הפתוחים בגריגה היה כ-3% בממוצע. היקף הנבירה הגבוה ביותר היה באביב והנמוך ביותר בקיץ (איור 4), וההבדלים ביניהם היו מובהקים (Friedman test, χ2=8.994, P=0.011). ניתוח זה אינו כולל את חורשות האורנים, שם היקף הנבירה היה גבוה ביותר בחורף, אך בקיץ ובאביב הנבירה הייתה נמוכה מאוד (איור 4). הבדלים אלה בין העונות היו מובהקים (Friedman test, χ2=22.435, P).
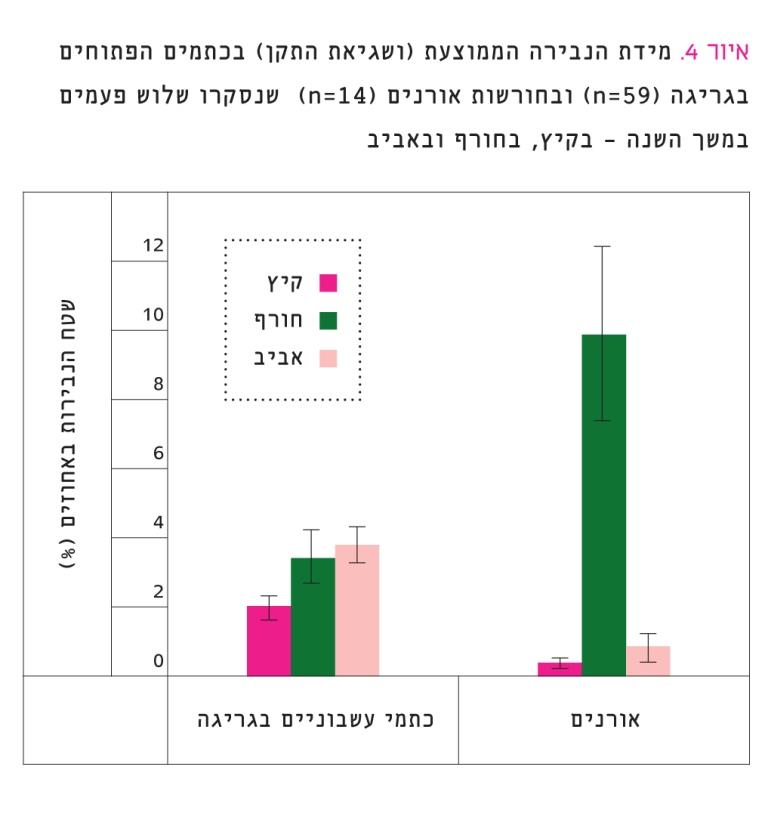
איור 4
מידת הנבירה הממוצעת (ושגיאת התקן) בכתמים הפתוחים בגריגה (59=n) ובחורשות אורנים (14=n) שנסקרו שלוש פעמים במשך השנה – בקיץ, בחורף ובאביב
דיון
מספר מחקרים הראו כי משטר הפרעה הנגרם על-ידי יונקים בינוניים וגדולים [10, 17, 18, 29], והפרות הקרקע שהם גורמים להן [3, 19, 26], מעצבים את חברת הצומח בבית הגידול .Jones ועמיתיו [15, 16] הגדירו אורגניזמים המשנים את מבנה בית הגידול ומעצבים בכך חברה שונה, כ‘מהנדסי סביבה‘. הם טענו שפעילותם של ‘מהנדסי הסביבה‘ משנה את פיזורם ואת זמינותם של משאבים במרחב וכך גם את תפוצתם של מינים שונים. חזירי בר הנוברים בקרקע גורמים להפרעה פיזית של הקרקע המשנה את האוורור בה, ויכולה להגדיל את מידת החדירות למים, להביא לקליטת זרעים גבוהה יותר וליצירת תנאים משופרים לנביטתם. כמו כן, הנבירה יכולה לפגוע בבנק הזרעים בקרקע [13].
נבירת קיץ הורידה מאוד את צפיפות הפרטים באביב העוקב לעומת חלקות שלא נברו בהן, אך לא שינתה את עושר המינים ואת הביומסה (איור 1ב). מגוון המינים בשטחים אלו היה גבוה מזה שבנבירת החורף ובחלקות הבקרה (איור 1א). לנבירת הקיץ הייתה השפעה שלילית ניכרת על השפע של המין הנפוץ ביותר, שבמרבית המקרים היה עוקצר מצוי (.Brachypodiumdistachyum L). נבירת הקיץ תפקדה כנראה גם כמלכודת זרעים. בחלקות אלה נמצאו 26 מינים ייחודיים שלא הופיעו כלל בחלקות האחרות, לעומת 14 מינים ייחודיים שנמצאו רק בחלקות ללא נבירה. מניתוח התוצאות עולה כי 12 מינים נפוצים היו קשורים לנבירה חזקה ועוד 5 לנבירה חלשה ובינונית (איור 3), וכולם הופיעו גם בחלקות שחזירי הבר נברו בהן בקיץ. הנבירה העניקה כנראה יתרון להתפתחות מינים אלה על-ידי דחיקה של מינים אחרים, אם על-ידי פגיעה בבנק הזרעים בקרקע או בצורה עקיפה על-ידי שינוי תנאי הסביבה הדרושים לנביטה ולהתבססות שלהם. במקביל נמצאו בניתוח התוצאות 7 מינים נפוצים שנפגעו מהנבירה ונמצאו בעיקר בחלקות ללא נבירה. מינים אלה, וככל הנראה גם חלק מהמינים הייחודים שנמצאו בחלקות ללא נבירה, נפגעו בעקבות השתנות תנאי טרום הנביטה ובנק הזרעים או בעקבות פגיעה ישירה בצמח עצמו לאחר הנביטה וחוסר יכולתו להתחדש.

נבירת החורף, למרות שבוצעה בעיצומה של עונת הצמיחה, לא שינתה את צפיפות הפרטים ואת עושר המינים יחסית לחלקות שלא נברו בהן. הצומח בחלקות אלה היה מורכב מפרטים ששרדו את הנבירה ומפרטים שנבטו מבנק הזרעים בקרקע לאחר הנבירה. עושר המינים בחלקות נבירת החורף היה נמוך מאשר בחלקות נבירת הקיץ. הצמחים העשבוניים בחלקות נבירת החורף הצליחו להתאושש לאחר הנבירה כפי שניתן לראות מנתוני הצפיפות ועושר המינים. עם זאת הם היו מוגבלים בצמיחתם בשארית תקופת הצימוח ולכן לא הספיקו למצות את מלוא פוטנציאל הצמיחה שלהם, כפי שניתן לראות בהשפעה השלילית על הביומסה (איור 1ב). בחלקות נבירת החורף נמצאו כ-12 מינים ייחודיים, פחות מאשר בנבירת הקיץ ובחלקות שלא נברו בהן. מינים שנטו להימצא לאחר נבירה חזקה הופיעו בחלקות נבירת החורף, אך לא כך לגבי המינים שנמצאו לאחר נבירה חלשה ובינונית, דבר שמחזק את הקשר של הראשונים לעצם קיום הנבירה. מינים אלה הצליחו להתבסס ולצמוח בשארית העונה בשטח שעבר נבירה חזקה בעוד שבחלקות שהייתה בהן נבירה קלה ובינונית השתקמה ברובה הצמחייה שנכחה שם לפני הנבירה. נבירות בעוצמות שונות, בהיקפים שונים ובעונות שונות יוצרות למעשה נישות התחדשות שיש בהן מגוון של תנאים הגורמים לשינוי בהרכב הצומח העשבוני.
היקף הנבירות שנעשו על-ידי חזירי הבר בשטחים הפתוחים בגריגה נשמר ברמה של 4%–2 במשך כל עונות השנה (איור 4), וזאת בדומה להיקף הנבירה במקומות אחרים בעלי אקלים ים תיכוני [6]. בקיץ ירד היקף הנבירה, כנראה בשל התייבשות ומיעוט האורגניזמים בחלק העליון של הקרקע. הירידה ברמת הפעילות בקיץ מוסברת גם על-ידי מעבר חזירי הבר לשטחים חקלאיים ולגינות נוי מושקות [6]. בחורשות אורנים היו מעט נבירות באביב ובקיץ. בחורף מאכלס מצע המחטים הרטוב פטריות וחסרי חוליות, ולכן היקף הנבירות בחורשות היה 10% מכלל השטח הניתן לנבירה (איור 4). תוצאות המחקר מראות כי חזירי הבר משנים את היקפי הנבירה שלהם בין עונות, תצורות צומח, ומפנים טופוגרפיים (איור 4 ונתונים שלא פורסמו). הם חוזרים לאתרים מועדפים שנברו בהם בעבר ובו בזמן מזהים אזורי שיחור חדשים ומשנים אתרי נבירה. אחוז הנבירה השנתי המצטבר בשטחים הניתנים לנבירה בגריגה הוא כ-9% ולכן ניתן להעריך כי ייתכן שבטווח של 10–11 שנים ינברו חזירי הבר בכל השטח הניתן לנבירה בגריגה.
היקף הנבירה הגבוה (בעיקר בקיץ) וההשפעה שיש לו על המגוון מלמדים על השפעת חזירי הבר על הקרקע ועל הצמחים. השפעה זו תומכת בהגדרתם כ‘מהנדסי סביבה‘ המשנים את התנאים בבית הגידול ואת חברת הצמחים העשבוניים. מנגד, לא מצאנו השפעה גדולה של הנבירה על חברת הצומח העשבוני בצורה המצדיקה הגדרה של שטחי הנבירות ככתם מבני חדש. הביומסה בחלקות נבירת הקיץ הייתה דומה לזו שבחלקות ללא הנבירות, למרות שמספר הפרטים היה רק 60% מזה שבחלקות הבקרה. לא מצאנו הבדלים בצפיפות הפרטים ובעושר המינים בין נבירת החורף לחלקות ללא נבירות, אבל הביומסה בהן הייתה שונה. יתרה מכך, על סמך תצפיות בשנה העוקבת, נראה שההשפעות שנמצאו הן קצרות מועד ואינן נמשכות יותר מעונה אחת, אולם נקודה זו דורשת מחקר המשך.
חזירי בר יוצרים משטר הפרה בהיקף נרחב, אך הפרה זו אינה משנה בצורה ניכרת את תנאי בית הגידול העונתי, וזאת בשונה ממינים המוכרים כ‘מהנדסי סביבה‘ מובהקים [15, 16, 29], כמו הדרבנים בנגב (Hystrixindica) המייצרים הפרה בקנה מידה קטן במרחב אך יוצרים כתם בעל ניגוד גדול לסביבה [3]. נראה כי הצומח העשבוני באזור הים תיכוני בישראל, שהתפתח במשך זמן אבולוציוני ארוך תחת השפעה של נבירה מצד חזירי בר מותאם למשטר הפרה זה כפי שהוא מותאם לרעייה [21]. לעומת זאת, במקומות שחזירי הבר הם מין פולש בהם נמצאו השפעות ברורות [9, 27] ושם הוגדרו חזירי בר כ‘מהנדסי סביבה‘ מובהקים [8].
תוצאות המחקר שלנו מצביעות אמנם על פעילות הפרה נרחבת של חזירי הבר אולם אין בפעילות זו כדי לגרום לפגיעה במגוון הצמחים העשבוניים, אלא להפך, הן מציגות את תרומתו של חזיר הבר לשימור המגוון של החברה העשבונית ברמת הנדיב. לפיכך הן מדגישות את החשיבות בניטור אוכלוסיות חזירי בר בשטחי שמורה ובמסדרונות בין שמורות, בטרם קבלת החלטות ממשקיות שישפיעו על חזירי הבר בעקבות נזקים לחקלאות וחדירה לאזורים מיושבים.
תודות
אנו רוצים להודות לרמת הנדיב על מימון המחקר. תודה מיוחדת לליאת הדר ולכל צוות רמת הנדיב על העזרה הרבה בביצוע המחקר.
השפעתם של חזירי הבר על שטחים חקלאיים ואף בתוך יישובים כפריים מוכרת כמזיקה. חזירי בר נוהגים לחפור ולנבור בקרקע במטרה לחפש מזון. פעילות נבירה זו מתקיימת גם בשטחים הטבעיים שחיים בהם חזירי הבר. כיונק היבשתי הטבעי הגדול בישראל, יש לחזירי הבר היכולת להפר את האיזון של המערכת האקולוגית ואף לשנותה. במחקר שנערך בפארק רמת הנדיב שבדרום הכרמל, נבחנה השפעת הנבירה בקרקע על הצמחייה העשבונית הים תיכונית (צמחים נמוכים שמחזור החיים שלהם קצר, ואינו עולה על שנה). נמצא שבעוד שהנבירה בולטת מאוד בנוף ואף משתרעת על פני שטחים נרחבים, הרי שהשפעתה האקולוגית על הצמחייה העשבונית הייתה קטנה בלבד, ואף נמצאה בה תועלת מבחינת שימור מגוון מיני הצומח העשבוני. ייתכן שהפער הזה נוצר מכיוון שמערכות טבעית כגון זו שבאזור הנבדק הספיקו לעבור תהליכי הסתגלות אִטיים (מבחינת הרכב המינים ויכולתם להתאושש) להפרעות שיוצרות נבירות חזירי הבר בקרקע, בדומה להשפעות של רעיית צאן ובקר. מסיבות אלה, חזירי הבר אינם יכולים להיחשב 'מהנדסי סביבה' באזור הים תיכוני. הנזקים שגורמים חזירי בר בעקבות חדירה לאזורים חקלאיים ומיושבים יוצרת לחץ לקבלת החלטות שיביאו לצמצום השפעתם, אך חשוב שתילקח בחשבון גם תרומתם בפועל לשימור מגוון המינים של מיני עשב. יש חשיבות לניטור של אוכלוסיות חזירי הבר ושל השפעתן על מערכות אקולוגיות – בשטחים פתוחים ובמסדרונות האקולוגיים שביניהם – כדי ליצור בסיס לקבלת החלטות מושכלת
מערכת אקולוגיה וסביבה
מקורות
- קפלן מ. 1988. קרקעות רמת הנדיב – דו“ח מחקר מס‘ 2. תל-אביב: החברה להגנת הטבע.
- Agra H and Ne’eman G. 2009. Woody species as landscape modulators: Their effect on the herbaceous plants in a Mediterranean maquis. Plant Ecology 205: 165-177.
- Boeken B, Shachak M, Gutterman Y, and Brand S. 1995. Patchiness and disturbance: Plant community responses to porcupine diggings in the central Negev. Ecography 18: 410-421.
- ter Braak CJF and Smilauer P. 2002. CANOCO reference manual and CanoDraw for Windows user’s guide version 4.5. Biometris – Plant Research International, Wageningen and Ceska Budejovice.
- ter Braak CJF and Smilauer P. 2003. CANOCO for Windows version 4.51. Biometris – Plant Research International, Wageningen.
- Cahill S, Llimona F, and Gracia J. 2003. Spacing and nocturnal activity of wild boar Sus scrofa in a Mediterranean metropolitan park. Wildlife Biology 9: 3-13.
- Cocca G, Sturaro E, Dal Compare L, and Ramanzin M. 2007. Wild boar (Sus scrofa) damages to mountain grassland. A case study in the Belluno province, eastern Italian Alps. Italian Journal of Animal Science 6: 845-847.
- Crooks JA. 2002. Characterizing ecosystem-level consequences of biological invasions: The role of ecosystem engineers. Oikos 97: 153-166.
- Cushman .H, Tierney TA, and Hinds JM. 2004. Variable effects of feral pig disturbances on native and exotic plants in a California grassland. Ecological Applications 14: 1746-1756.
- Dublin HT, Sinclair ARE, and Mc Glade J. 1990. Elephants and fire as causes of multiple stable states in the Serengeti-Mara woodlands. The Journal of Animal Ecology 59: 1147-1164.
- Engeman RM, Smith HT, Severson R, Severson MA, Shwiff SA, Constantin B, and Griffin D. 2004. The amount and economic cost of feral swine damage to the last remnant of a basin marsh system in Florida. Journal for Nature Conservation 12: 143-147.
- Herrero J, Serrano AG, Cuto S, Ortuno VM, and Gonzalez RG. 2006. Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. European Journal of Wildlife Research 52: 245-250.
- Hyatt LA. 1999. Differences between seed bank composition and field recruitment in a temperate zone deciduous forest. The American Midland Naturalist 142: 31-38.
- Ickes K, Dewalt SJ, and Appanah S. 2001. Effects of native pigs (Sus scrofa) on woody understorey vegetation in a Malaysian lowland rain forest. Journal of Tropical Ecology 17: 191-206.
- Jones CG, Lawton JH, and Shachak M. 1994. Organisms as ecosystem engineers. Oikos 689: 373-386.
- Jones CG, Lawton JH, and Shachak M. 1997. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78:1946-1957.
- Knapp AK, Blair JM, Briggs JM, Collins SL, Hartnett DC, Johnson LC, and Towne EG. 1999. The keystone role of Bison in North American tallgrass prairie. BioScience 49: 39-50.
- McInnes PF, Naiman RJ, Pastor J, and Cohen Y. 1992. Effects of moose browsing on vegetation and litter of the boreal forest, Isle Royale, Michigan, USA. Ecology 73: 2059-2075.
- Miller B and Ceballos G. 1994. The prairie dog and biotic diversity. Conservation Biology 8: 677-681.
- Moody A and Jones JA. 2000. Soil response to canopy position and feral pig disturbance beneath Quercus agrifolia on Santa Cruz Island, California. Applied Soil Ecology 14: 269-281.
- Perevolotsky A and Seligman NG. 1998. Degradation of Mediterranean rangeland ecosystems by grazing: Inversion of a paradigm. BioScience 48: 1007-1017.
- Rixen C, Haag S, Kulakowski D, and Bebi P. 2007. Natural avalanche disturbance shapes plant diversity and species composition in sub-alpine forest belt. Journal of Vegetation Science 18: 735-742.
- Royo AA and Carson WP. 2005. The herb community of a tropical forest in central Panama: Dynamics and impact of mammalian herbivores. Oecologia 145: 66-75.
- Shachak M, Boeken B, Gruner E, Kadmon R, Lubin Y, Meron E, Ne’eman G, Perevolotsky A, and Shkedy Y. 2008. Woody species as landscape modulators and their effect on biodiversity patterns. BioScience 58: 209-221.
- Schley L and Roper T J. 2003. Diet of wild boar Sus scrofa in western Europe, with particular reference to consumption of agricultural crops. Mammal Review 33: 43-56.
- Tardiff SE and Stanford JA. 1998. Grizzly bear digging: Effects on subalpine meadow plants in relation to mineral nitrogen availability. Ecology 79: 2219-2228.
- Tierney TA and Cushman JH. 2006. Temporal changes in native and exotic vegetation and soil characteristics following disturbances by feral pigs in a California grassland. Biological Invasions 8: 1073-1089.
- Welander J. 2000. Spatial and temporal dynamics of wild boar (Sus scrofa) rooting in a mosaic landscape. Journal of Zoology 252: 263-271.
- Wright JP, Jones CG, and Flecker AS. 2002. An ecosystem engineer, the beaver, increases species richness at the landscape scale. Oecologia 132: 96-101.


 מהדורה מודפסת
מהדורה מודפסת








