
חזית המחקר
השפעתה של נמלת הקציר על הקרקע והצמחייה בצפון הנגב
סטאפן לואי המכונים לחקר המדבר, ע"ש יעקב בלאושטיין, אוניברסיטת בן-גוריון בנגב
פועה בר (קותיאל) המחלקה לגאוגרפיה ופיתוח סביבתי, אוניברסיטת בן-גוריון בנגב
אריה בודובסקי מו"פ יהודה

יכולת ההִנדוס של נמלת הקציר מתבטאת בהגדלת השטח היחסי שמתפקד כמשמר מים, בהעלאת פוריות הקרקע ויצרנותה, וכפועל יוצא מכך – בזירוז ההשתקמות של שטח מופר | צילום: עמיר מור-מוסרי
מאת
עמיר מור-מוסרי
המחלקה לגאוגרפיה ופיתוח סביבתי, אוניברסיטת בן-גוריון בנגבסטאפן לואי
המכונים לחקר המדבר, ע"ש יעקב בלאושטיין, אוניברסיטת בן-גוריון בנגבפועה בר (קותיאל)
המחלקה לגאוגרפיה ופיתוח סביבתי, אוניברסיטת בן-גוריון בנגבאריה בודובסקי
מו"פ יהודהמאת
עמיר מור-מוסרי
המחלקה לגאוגרפיה ופיתוח סביבתי, אוניברסיטת בן-גוריון בנגבסטאפן לואי
המכונים לחקר המדבר, ע"ש יעקב בלאושטיין, אוניברסיטת בן-גוריון בנגבפועה בר (קותיאל)
המחלקה לגאוגרפיה ופיתוח סביבתי, אוניברסיטת בן-גוריון בנגבאריה בודובסקי
מו"פ יהודהלממשק של מערכת אקולוגית שאינו תואם לתנאי הסביבה, כגון רעיית יתר באזור צחיח, יש השפעות הרסניות על המערכת לטווח הזמן הקצר והארוך. שיקום יעיל בטווח זמן קצר יחסית דורש התערבות של האדם על-ידי שילוב אורגניזם בעל יכולת "הנדוס סביבתי" יחד עם עידוד של ריבוי או התפשטות של המינים המקומיים. מרבית המחקרים באזורים צחיחים התמקדו בעיקר בשיחים כמהנדסי סביבה ופחות בהשפעת אורגניזמים אחרים, כמו חרקים, על הסביבה. במחקר הנוכחי השווינו את השפעת קִני שני מינים של נמלת הקציר, השחורה Messor ebeninus והחולית – Messor arenarius, על כתמי נוף בשטחי שיחים במדבר (בתת ספר ערבתית). השטחים שנבדקו נמצאים באזור חירן (מערבית לערד, בצפון מזרח הנגב), בשטח שיש בו רעיית יתר ובשטח בעל רעייה מתונה שהיה חשוף בעברו לרעיית יתר. תוצאות המחקר מראות שלנמלי הקציר יש השפעה משמעותית על פוריות הקרקע ועל הצמחייה, שניכרה לא רק בשנה גשומה אלא אף בשנת בצורת. תוצאות המחקר מצביעות על הפוטנציאל של נמלים ככלי ממשק לשיקום שטחי מדבר פגועים.
על קצה המזלג
- לרעיית יתר יש השפעה שלילית על המערכת האקולוגית בנגב ובאזורי מדבר אחרים בעולם.
- נמלי הקציר כ’מהנדסות סביבה’ עשויות לסייע בשיקום שטחי שיחים בתהליכי התדרדרות.
- התיישבות נמלים יוצרת כתמים עשירים במשאבים, המגדילים את הייצור הראשוני של ביומסת העשבוניים.
- יחסי המשוב בין שיחים לנמלים מגדילים את פוריות המערכת האקולוגית.
- לנמלי הקציר פוטנציאל לתרום לשיקום שטחים מדבריים פגועים.
המערכת
הלכה למעשה
מבוא
לרעיית יתר יש השפעה שלילית על המערכת האקולוגית בנגב [28] ובאזורי מדבר אחרים בעולם [4]. השפעה זאת מתבטאת בירידה במגוון מיני הצומח והחי [23], בפגיעה בתכונות הקרקע [16], בשינוי המאפיינים של יחידות שטח קטנות בעלות מאפיינים מיקרו-טופוגרפיים או צמחים אופייניים המובדלות מסביבתן מבחינת מבנה, גודל ופיזור השונים בשטח נתון [37] ואף בפגיעה במאפיינים המָקרו-טופוגרפיים של השטח [1]. במצבים קיצוניים – כמו באזורים נרחבים בצפון הנגב [28] – שיקום השטח אינו יכול להסתמך על תהליכים טבעיים, ומצריך את התערבות האדם על-ידי שינוי שיטת הממשק [2].
השיטה המקובלת לשיקום שטח שנפגע מרעיית יתר מבוססת על תיחום השטח הפגוע ומניעת רעייה למשך מספר שנים [6]. שיטה זו בעייתית מכמה סיבות: הזמן הרב הנדרש לשיקום, עלותה הגבוהה והתלות הרבה של התושבים באזורים אלה ברעייה כמקור פרנסה [36]. גישה משלימה המאפשרת האצה של תהליכי השיקום הטבעיים מבוססת על שימוש ב"מהנדסי סביבה" (ecosystem engineer, אורגניזם שמשנה, יוצר או הורס בית גידול, ומשפיע ישירות או בעקיפין על זמינות משאבים למינים אחרים על-ידי יצירת שינוי פיזיקלי בגורמים ביוטיים ואביוטיים [19]).
מושג זה נחקר בעיקר בהקשר של צמחים (בדגש על בני שיח) [18, 31], ובמידה מועטה בהקשר של אורגניזמים מקבוצות טקסונומיות אחרות (כגון טחבנים [34], דורבנים [45] ועדרי צאן [16, 37]). תצפיות מקדימות שערכנו בצפון הנגב בשטחים החשופים לרעיית יתר בשנים עוקבות הראו היעדר התבססות חדשה של בני שיח, בעוד שמספר קִני הנמלים וגודלם (הנטושים והפעילים) נשארו קבועים, ובחלק גדול של השטחים אף נצפתה עלייה בגודל הקִנים ובכיסוי שלהם. לשם בחינה וכימות של השפעת קִני נמלת הקציר (השחורה – Messor ebeninus והחולית – Messor arenarius [38]) על המערכת האקולוגית בצפון הנגב נבדקו מספר מדדים אקולוגיים בקִנים, בכתמי השיח ובתווך הפתוח שבין הקִנים ובני השיח. המדדים שנבדקו היו ביומסת עשבוניים, גודל הקִנים ומספרם [23], תאחיזת מים וריכוז חומר אורגני וחומרי הזנה (nutrients) בקרקע.
בשטח המחקר נצפו גם פרטים מהמין נווטת כתומה (Cataglyphis lividus) ,אך עקב נדירותו היחסית למינים נמלת הקציר השחורה והחולית, אין התייחסות אליו במאמר זה [42].
מטרות המחקר היו לבדוק את השפע והפיזור של קִני נמלת הקציר בבתת ספר ערבתית, לכמת את השפעת הקִנים על המערכת האקולוגית במונחים של תפרוסת בשטח, פוריות ויצרנות של צמחים עונתיים, ולהעריך את פעילותן כמהנדסי סביבה באזורים עם רעיית יתר ותחת רעייה מתונה.

שיטות
שטח המחקר
המחקר נעשה באזור חירן (31o19'34"N, 34o59'04"E), מצפון ליישוב הבדואי חורה ומדרום ליער יתיר בצפון הנגב. האזור גבעי (גובה עד 250 מ' מעל פני הים) והקרקע חולית-חרסיתית [41]. האקלים מדברי: כמות הגשם השנתית הממוצעת נעה בין 150 ל-200 מ"מ, הטמפרטורה בעונה הקרה (דצמבר–פברואר) 8.1–19.2 מעלות צלזיוס, ובעונה החמה (יוני עד אוגוסט) 21–34 מעלות (נתוני השירות המטאורולוגי).
המחקר המוצג הוא חלק מלימוד ארוך טווח על בתות ספר ערבתיות באזור בין 2006 ל-2014, שכלל תצפיות של הכותבים ובעלי החוות. המדידות הקשורות למחקר המוצג לעיל נערכו בעיקרן בין 2008 ו-2010. שנת 2009 מייצגת שנת בצורת בהשוואה ל-2008, למרות כמויות הגשם הדומות – 155 ו-159 מ"מ לשנה בהתאמה – מאחר שהיא התאפיינה בפיזור גשמים לא אחיד לאורך העונה, דבר שהתבטא בירידת מרבית כמויות הגשם בתחילת העונה ובסופה, לעומת הפיזור האחיד יחסית בשנת 2008. לאורך עונת הגשמים ב-2010 נמדדו 237 מ"מ (נתוני השירות המטאורולוגי).
מבחינת הצומח האזור מוגדר כבתת ספר ערבתית [14, 37], שהייתה נתונה עשרות שנים לרעיית יתר לא מבוקרת עד 1992. הדבר מתבטא בכיסוי צמחי נמוך (60–70 בני שיח לדונם), ובעקבות זאת אף בסחף קרקע ובהיווצרות חריצים (rills) וערוצים מתחתרים (gullies) [23, 1]. בשנים 1992–1994 הוקמו באזור מספר חוות בודדים למטרות שימור הקרקע מפני סחיפה והעלאת פוריותה על בסיס של קיום רעייה בת-קיימא (בהתאם לעקרונות של Swanson) [39]. לאחר הקמתן נצפה בשטחי הרעייה של חוות אלה שיקום הדרגתי בפוריות הקרקע, בעוד שבשטחים בסביבתן המשיכה רעיית יתר מלווה בירידה משמעותית של כסות הצומח [26] ובהאצת סחף הקרקע.
לצורך בחינת השפעתן של הנמלים על הקרקע והצומח בשטחים החשופים לרעיית יתר ותחת רעייה מתונה, נבחרו שני שטחי בתה מייצגים בגודל 15X15 מ', שכ-25% מהם מכוסים סלעים, כולם בעלי מפנה דרומי ומגוון צמחים דומה, בהם בני שיח בעיקר ממוצא אירנו-טורני (ערבתיים) וסהרו-ערביים ונציגים בודדים של המין סירה קוצנית (ממוצא ים תיכוני). שטח אחד התאפיין ברעייה מתונה ונמצא בתחומי "חוות יתיר", והשטח השני נמצא 200 מ' מזרחית לו מחוץ לתחומי החוות, באזור שהתאפיין ברעיית יתר.
הגדרת השטח כמתאפיין ברעיית יתר נעשתה על סמך תצפיות ומדידות רבות שונות באזור בטרם המחקר ובהתאם לשיחות שנערכו עם בעלי החוות באזור, שהושוו לתיאור של Olsvig-Whittaker ואחרים [28].
תצפיות נוספות נאספו בשנת 2014 מחלקות משטח מיזם ואדי עתיר, המצוי 200 מ' מדרום לחורה, 7 ק"מ דרום-מזרחית לשטח המחקר בחוות יתיר. השטח סבל מסחף קרקע רב, ושוקם בחלקו ב-2012 על-ידי הקמת לימנים (שטחים נמוכים ושטוחים לאגירת המים והסחף) [35]. חלק ממנו נשמר מעיבוד וחלק נזרע בחיטה [27]. צמידות יחידות השטח תחת הניהול השונה אפשרה בחינה עקבית של התנחלות נמלי הקציר באזורי מדבר מופָרים החל בהפרה ועד לשיקום.

הגדרת כתמי הנוף
אזורי בתת ספר ערבתית מתאפיינים בכתמי נוף [14], כלומר ביחידות שטח קטנות בעלות מאפיינים מיקרו-טופוגרפיים או צמחים אופייניים, המובדלות מסביבתן. בתצפיות בשנים הקודמות למחקר בשטחים המקיפים נוכחנו בדמיון הרב בין המאפיינים המיקרו-טופוגרפיים של כתמים בנוף המכילים קִני נמלים לאלה המכילים צומח רב-שנתי, ובשונות הקיימת בין המאפיינים של שני סוגי הכתמים הללו לבין אלה של התווך המקיף. מסיבה זו נבדקו השפעת הקִנים והשפעת הצומח הרב-שנתי באותם כלים [38]. לשם בחינת כתמי הנוף השונים והשוואתם לסביבה המקיפה הוגדרו בשני שטחי המחקר (רעייה מתונה ורעיית יתר) מאפייני הכתמים על בסיס הנקודות הבאות:
- שטח תחוח עם מאפיינים מיקרו-טופוגרפיים בולטים ומובדלים מהסביבה (מוגבהים או מונמכים).
- שטח ללא הפרות קרקע כתוצאה מפעילות אדם או בעלי חיים (חפירות מכרסמים וכדומה) מלבד פעילות נמלים.
- שטח המכיל קן נמלים או צמחים רב-שנתיים (שיחים, בני שיח ועשבוניים רב-שנתיים – גאופיטים והמיקריפטופיטים) או צמחים חד-שנתיים. משום גודלם של הצמחים החד-שנתיים ונופם הירוק בתחילתו של הקיץ הנחנו שיש להם השפעה דומה לזאת של הצמחים הרב-שנתיים על סביבתם [21].
האורגניזמים בכתמי הנוף חולקו לשלוש קבוצות בהתאם לסיווגם הביולוגי: קִני נמלים, להלן קִנים; שיחים ובני שיח, להלן בני שיח (כללו בעיקר: מתנן שעיר [Thyemeleae hirsuta], קזוח תלת-קרני [Pituranthos triradiatus], נואית קוצנית [Noaea mucronata] וסירה קוצנית [Sarcopoterium spinosum]) עשבוניים רב-שנתיים (עירית גדולה [Asphodelus ramosus], קיפודן מצוי [Echinops adenocaulus] ודרדר קרומי [Centaurea hyalolepis]).
נוסף על כך הוגדר התווך שבין כתמי הנוף. יש לציין, שחלק מכתמי הנוף הכילו מספר קבוצות התייחסות.
השפעת הקִנים על המאפיינים המיקרו-טופוגרפיים של בתת הספר הערבתית ועל מספר קבוצות ההתייחסות בתחומם
בשלב ראשון, נמדדו הציר האורכי והרוחבי של כתם הנוף, ושטחו חושב על-ידי הצבה במשוואת האליפסה [25]. הערכת גודל הקִנים התבססה על השטח שהפֵרה פעילות הנמלים (מיקומם של חורי כניסה, שפכי קרקע ונשר וכד').
נתוני כתם שהכיל יותר מקבוצת התייחסות אחת הוכנסו לניתוח נתוני כל אחת מהקבוצות הבונות את הכתם. לדוגמה, בכתם נוף בעל צירים של 0.5 ו-0.6 מ' (אורכי ורוחבי), שהכיל קן ושני שיחי מתנן, שויך לקִנים ולבני שיח שטח של 0.24 מ"ר (πX0.5/2X0.6/2). במסגרת ניתוח הנתונים לא הייתה התייחסות למספר הפרטים מאותה קבוצת התייחסות, אלא רק לנוכחותה בכתם הנוף. ממוצע של הערכים המשויכים לכל קבוצת התייחסות ייצג את השפעתה על הסביבה, בהנחה שככל שערך זה גדול יותר, כך השפעתה של קבוצת ההתייחסות על הסביבה רבה יותר [26]. ניתוח נתונים זה אפשר להעריך מדד נוסף המבטא את תמיכתה של כל קבוצת התייחסות בהטרוגניות של כתמי הנוף שהיא נמצאת בהם. מדד זה חושב על-ידי בחינת מספר קבוצות ההתייחסות בכל כתם נוף ושיוך מספר זה לקבוצות השונות שנמצאו (מדד שהוגדר כ-'הטרוגניות'). לדוגמה, בקן שמצוי בו דרדר קרומי שויך ערך הטרוגניות של 2 לקבוצות הקִנים והעשבוניים הרב-שנתיים, בעוד שלבן שיח בעל מופע יחידאי בכתם נוף נבדק, ניתן ערך הטרוגניות של 1. בשלב הבא חושבו לכל קבוצת התייחסות מספר המופעים שערך ה'הטרוגניות' שלהם שווה 2 או גדול ממנו, והם יכונו להלן מופעים 'מורכבים'.
התוצאה ניתנה כאחוז של כתמי הנוף ה'מורכבים' מכלל המופעים של קבוצת ההתייחסות.
דגימת הביומסה של הצמחים העונתיים
המדד המאפיין ביותר לזיהוי תהליכי מִדבור או תהליכי שיקום באזורי מדבר, הוא כושר הייצור הצמחי, בעיקר הביומסה של הצמחים העונתיים [23]. דגימות אקראיות של צמחים עשבוניים מהנוף נקצרו במרץ 2010 מתוך מלבנים בגודל 30X20 ס"מ משטחי הקִנים וכתמי בני השיח (כתמי נוף שהופעתן של שתי קבוצות ייחוס אלה בהם הייתה יחידאית) ומהתווך [32], בסך הכול 15 דגימות (חמש מכל יחידת נוף). הדגימות יובשו ב-60 מעלות למשך 48 שעות ונשקלו. המשקל בוטא בק"ג למ"ר.
דגימות הצמחים והקרקע נלקחו משולי הקִנים כדי למזער את הפגיעה בהם [43]. כמו כן, במחקר זה לא הייתה הבחנה בין קִני נמלים מאוכלסים ונטושים, מתוך הנחה שקיימות השפעות זהות על הסביבה בשני המקרים [13].
מלבד השוואת ממוצעי הביומסה של הצמחים העונתיים עבור כל יחידת נוף, נעשה שימוש בערכים שהתקבלו לחישוב התרומה של קִני הנמלים ובני השיח לכלל בתת הספר על-ידי כפל באחוז כיסויים מכלל השטח. לדוגמה, תרומת בני השיח לביומסה העונתית בשטח המחקר (Shrubs[HerbBM.Donation]) חושבה בצורה זו (הערכים מבוטאים בק"ג לדונם):
Shrubs(HerbBM.Donation) = (Shrubs(HerbBM) – Matrix(HerbBM) X Shrubs(Cover) X 1000)
הפחתת ממוצע הביומסה העונתית בכתמי השיח מהממוצע שהתקבל בתווך: ערכי ה-Shrubs(HerbBM) וה-Matrix(HerbBM) בהתאמה (ריכוזים בק"ג למ"ר).
הכפלה באחוז כיסוי הכתמים המכילים בני שיח מכלל הכתמים Shrubs(Cover).
הכפלה ב-1,000 לייצוג התוצאה עבור שטח של דונם (מ"ר).
דגימה ובדיקות קרקע
באפריל 2009 נאספו בשטח הרעייה המתונה שלוש דגימות מכל יחידת נוף. הדגימות נאספו מ-20 הס"מ העליונים של הקרקע, המייצגים את בית השורשים של מרבית הצמחים החד-שנתיים [26]. בסך הכול נאספו 12 דגימות קרקע בנות כחצי ליטר מהתווך, משולי הקִנים ומכתמי בני השיח (כתמי נוף שהופעתן של שתי קבוצות ייחוס אלה בהם הייתה יחידאית). הדגימות הועברו סמוך למועד הדגימה למעבדות לשירות שדה בגילת לבדיקות ריכוזי חומרי הזנה בעלי חשיבות למערכת האקולוגית המדברית (פחמן אורגני [11], חנקן כללי [5] וזרחן זמין P-PO3 [47]) ולאגירת המים בקרקע – קיבול שדה (field capacity [30]).
מלבד השוואת ממוצעי מדדים אלה בין יחידות הנוף נעשה שימוש בערכים המתקבלים לחישוב התרומה של הקִנים ובני השיח לכלל בתת הספר על-ידי כפל באחוז כיסויים מכלל השטח.
לדוגמה, תרומת הקִנים להעלאת כמות הזרחן בשטח המחקר (Nest[P.Donation]) חושבה בצורה זו (גרם לדונם):
Nest(P.Donation) = ([Nests(P) – Matrix(P)] X Nests(Cover)) X 0.2 X 1200 X103/103
בסיס הנוסחה: הפחתת ממוצע ריכוז הזרחן בקִני הנמלים מהממוצע שהתקבל בתווך – ערכי Nest(P) ו-Matrix(P) בהתאמה (ריכוזים במ"ג לק"ג קרקע) ולאחר הכפלה באחוז כיסוי הקִנים מכלל השטח Nests(Cover).
התאמת יחידות: הכפלה ב-0.2 (עומק בית השורשים של העונתיים, שבו צפויה עיקר התרומה של קבוצות הייחוס לפוריות בבתת הספר הערבתית, יחידות במ'). הכפלה ב-1,200 משקל סגולי של אדמה יבשה (ק"ג למ"ק). הכפלה ב-103, לייצוג התוצאה עבור שטח של דונם (יח' מ"ר). חלוקה ב-103, לייצוג התוצאה בגרמים.
המשקל הסגולי של הקרקע חושב כממוצע משקלים לאחר ייבוש של חמש דגימות קרקע של חצי ליטר ב-100 מעלות למשך 12 שעות [32].
ניתוח סטטיסטי
לבחינת המובהקות הסטטיסטית להבדלים בין התוצאות שהתקבלו ביחידות הנוף השונות בוצע מבחן ANOVA, α=0.05 בתוכנה JMP® ver. 5. לשם ניתוח הקשר בין מדדים שונים ומקדם הרגרסיה (r2) בוצע שימוש ב-EXCEL® ver. 10.

תוצאות
השפעת הקִנים על המאפיינים המיקרו-טופוגרפיים של בתת הספר הערבתית ועל קבוצות ההתייחסות האחרות
כבר בתצפיות המקדימות למחקר ניתן היה להבחין בתפוצה הרבה של הקִנים בשטח הרעייה המתונה בהשוואה ליתר כתמי הנוף. אישוש לתצפיות אלה מוצג באיור 1, המדגים תפרוסת הגבוהה ב-50% של הקִנים מבני השיח בשטח זה (40% ו-25% בהתאמה, איור 1א) וכן את גודלם הרב בהשוואה לכתמי הנוף המכילים את קבוצת הייחוס עשבוניים רב-שנתיים ב-2008 (ב-2009, שנת הבצורת, התקבלו ערכים דומים, איור 1ב). נוסף על כך, ניתן להבחין שבשטח רעיית היתר נמצאה תפרוסת יחסית דומה של כתמי השיח ושל קִני הנמלים. בהשוואה בין השנים השונות, נמצא שקיימת ירידה מובהקת בגודלי כתמי השיח וקִני הנמלים בין השנים 2008 ו-2009 (איור 1ב) בשטח הרעייה המתונה. בשטח רעיית היתר שמרו שתי קבוצות הכתמים על גודלם הקטן בין השנים השונות (איור 1ג).
באיור 1ד ניתן רמז נוסף לתרומתן של הנמלים למגוון האקולוגי בתחומי קִניהן, בכך ש-75% ממופעיהן היו מורכבים (כלומר, נמצאו קבוצות ייחוס נוספות), בעוד שאצל כתמי בני השיח המופעים המורכבים היו 25%.
קשר חיובי נמצא בין גודל קִני הנמלים לבין חופת חלק ממיני השיחים, כגון מתנן שעיר (איור 1ה) ומספר בני השיח שבתוכם (איור 1ו)
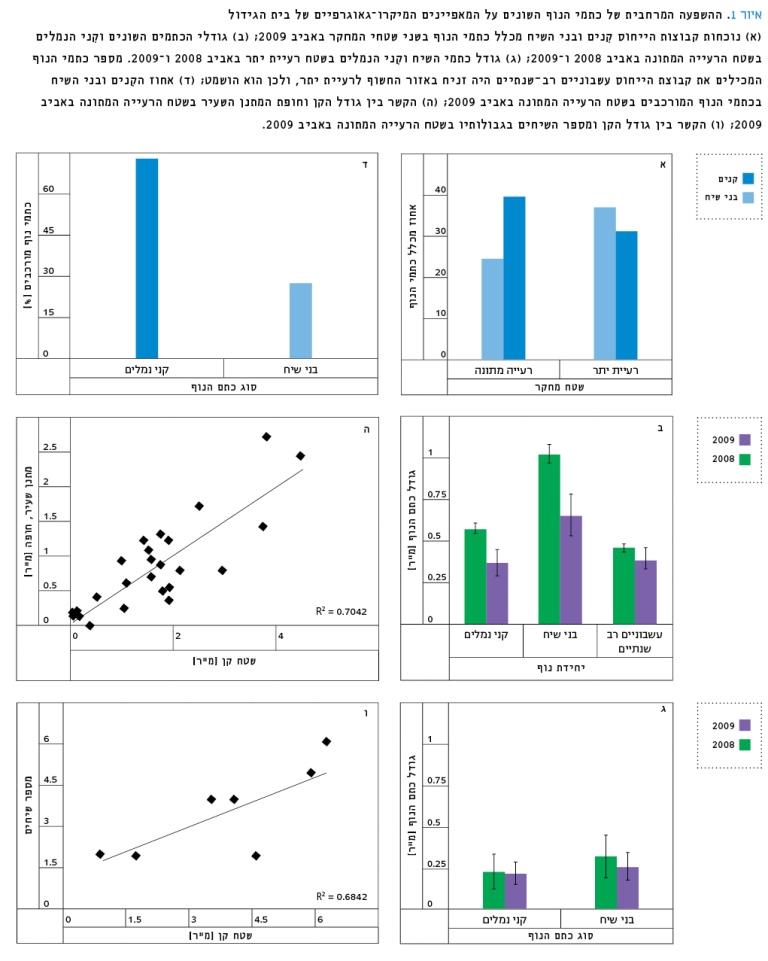
איור 1
ההשפעה המרחבית של כתמי הנוף השונים על המאפיינים המיקרו-גאוגרפיים של בית הגידול
א. נוכחות קבוצות הייחוס קִנים ובני השיח מכלל כתמי הנוף בשני שטחי המחקר באביב 2009; ב. גודלי הכתמים השונים וקִני הנמלים בשטח הרעייה המתונה באביב 2008 ו-2009; ג. גודל כתמי השיח וקִני הנמלים בשטח רעיית יתר באביב 2008 ו-2009. מספר כתמי הנוף המכילים את קבוצת הייחוס עשבוניים רב-שנתיים היה זניח באזור החשוף לרעיית יתר, ולכן הוא הושמט; ד. אחוז הקִנים ובני השיח בכתמי הנוף המורכבים בשטח הרעייה המתונה באביב 2009; ה. הקשר בין גודל הקן וחופת המתנן השעיר בשטח הרעייה המתונה באביב 2009; ו. הקשר בין גודל הקן ומספר השיחים בגבולותיו בשטח הרעייה המתונה באביב 2009.
מאפייני הצומח העונתי בכתמי הנוף השונים
בהערכה חזותית נמצא שבניגוד למגוון מיני העונתיים בכתמי בני השיח והעשבוניים הרב-שנתיים, קִני הנמלים התאפיינו בעיקר במינים ממשפחת הדגניים (Gramineae), כדוגמת מלעניאל מצוי (Stipa capensis), בן חיטה מעובה (Aegilops crassa) ובן חיטה מדברי (Aegilops Kotschyi), בעוד שביתר הכתמים רק כשליש מהחד-שנתיים היו דגניים. נוסף לשוני בהרכב הצמחייה, נמצא שהצמחים בקִני הנמלים (הפעילים והנטושים) גדולים ומפותחים יותר מאלה שבכתמים האחרים (איור 2).

איור 2
דוגמאות ליחידות הנוף המיקרו-גאוגרפיות שהוגדרו במחקר
א. קן נמלים בשטח הרעייה המתונה; ב. קן נמלים בשטח תחת רעיית יתר; ג. כתם נוף עם קבוצת הייחוס של בני השיח; ד. התווך שבין הכתמים. רוחב צלע ריבוע הברזל 0.5 מ'.
בחישוב כלל הביומסה נמצא שבקִני נמלים יש כמות גבוהה פי שישה (ובמובהקות סטטיסטית של α=0.05) בהשוואה לתווך ולכתמי הנוף המכילים את בני השיח (איור 3).
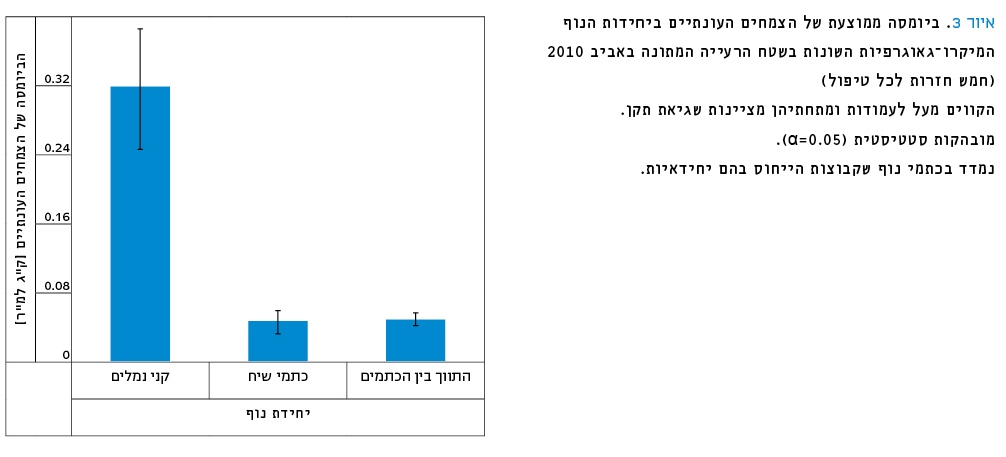
איור 3
ביומסה ממוצעת של הצמחים העונתיים ביחידות הנוף המיקרו-גאוגרפיות השונות בשטח הרעייה המתונה באביב 2010 (חמש חזרות לכל טיפול).
הקווים מעל לעמודות ומתחתיהן מציינות שגיאת תקן.
*מובהקות סטטיסטית (α=0.05)
נמדד בכתמי נוף שקבוצות הייחוס בהם יחידאיות.
השפעת הקִנים על הקרקע של בתת הספר הערבתית
באיור 4א ניתן לראות כי לקִנים ערך קיבול שדה הנמוך במעט בהשוואה לכתמי הנוף המכילים את בני שיח וגבוה בכ-5% מזה המצוי בתווך. בקִנים נמצא שהחומר האורגני, הזרחן וכמות החנקן הכללי באביב היו גבוהים מהערכים שנמצאו בתווך, ושווים למה שנמצא בכתמי הנוף של בני השיח או גבוהים ממנו במקרה של הזרחן (איור 4 ב–ד).
למרות ההבדלים הניכרים, לא נמצאה מובהקות סטטיסטית (α=0.05), דבר המוסבר במספר הדגימות המועט שנבע ממגבלות המחקר.
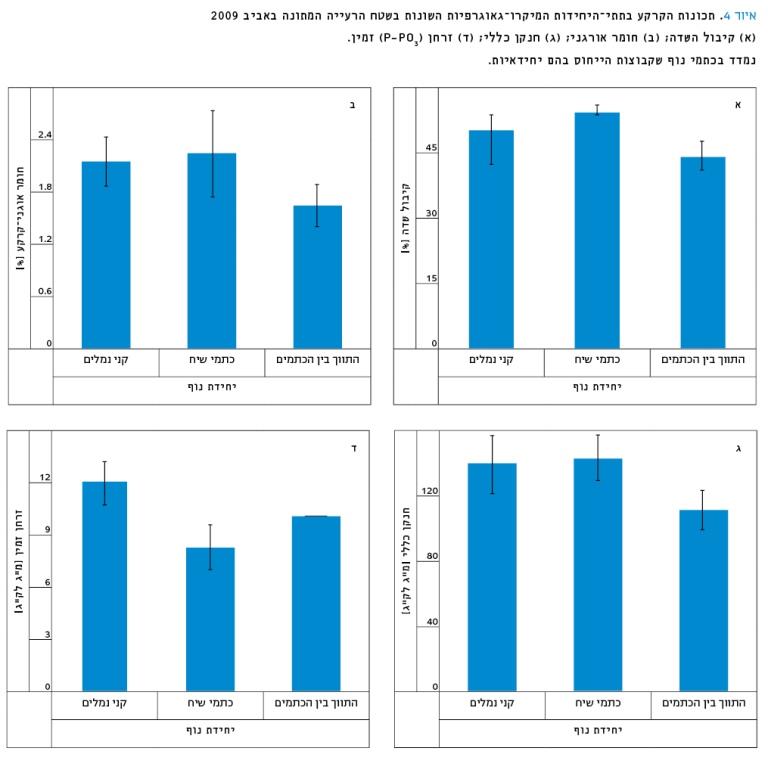
איור 4
תכונות הקרקע בתתי-היחידות המיקרו-גאוגרפיות השונות בשטח הרעייה המתונה באביב 2009
א. קיבול השדה; ב. חומר אורגני; ג. חנקן כללי; ד. זרחן (P-PO3) זמין.
נמדד בכתמי נוף שקבוצות הייחוס בהם יחידאיות
דיון
מטרתו העיקרית של המחקר הייתה להעריך את יכולת ההנדוס הסביבתי של קִני נמלים בשטחי שיחים בצפון הנגב [7, 10]. נוסף על השפעת הקִנים על סביבתם הפנימית המתוארת במחקרים אחרים [6, 7], ועל היותם מדדים ביולוגיים לפוריות [8], נמצא שיש להם השפעה של ממש על הסביבה המקיפה אותם. כימות ההיבט המרחבי-אקולוגי בוצע על-ידי מדידות גודל ומספר של קִני הנמלים בשטח החשוף לרעיית יתר ובשטח תחת רעייה מתונה (שבעברו היה חשוף לרעיית יתר). הנחנו שההבדל בין השטחים הללו יצביע על הדינמיקה של הנמלים, מבחינת התבססותן והרחבת קניהן במעבר ממצב של רעיית יתר לניהול השטח על בסיס רעייה מתונה, כמתואר בספרות לגבי בני שיח [29]. ההשוואה בין השנים שונות יכולה לרמז על הדינמיקה תלוית האקלים, וההשוואה לכתמי בני השיח נותנת נקודות ייחוס לדינמיקה זאת. ניתן להבחין שבשטח החשוף לרעיית יתר מספר הקִנים היה דומה לזה שבכתמי בני השיח, אך לאחר הורדת לחץ הרעייה חלה התרבות מהירה של קִני הנמלים (שתרמה להתרבות העשבוניים בשטחי הקִנים). נוסף על כך, עמידותם של קִני הנמלים לתנאי הסביבה ניכרת בשמירה על גודלם ועל מספרם בשטח הנתון לרעיית יתר אף בשנת בצורת, כמו זו שהייתה ב-2009 (בהשוואה ל-2008) (איור 1ב), ובירידה מתונה בגודל הקִנים בשטח שנתון לרעייה מתונה (איור 1ג). הערכת גודל הקִנים התבססה על הפָרות הקרקע החיצוניות של הנמלים (שפכי קרקע [nests' soil dumps], שפכי שלף וחורי כניסות) ולא לפי סך השטח שתופסים התעלות וחדרי הקן, שמוערך כגדול בהרבה [40], כך שסביר שההשפעה על הסביבה רחבה יותר מהמתואר לעיל. קשרים אקולוגיים נוספים בין קִני הנמלים לסביבה הצמחית אותרו בין גודל קִני הנמלים למספר שיחי המתנן ולגודל חופתם. אף על פי שהקשר המתואר אינו מצביע ישירות על קִני הנמלים כגורם המשפיע על שיחי המתנן, התצפיות המתוארות ואחרות שנאספו מאוחר יותר מאזורים אחרים, מצביעות על קִני הנמלים כגורם המשפיע על הצמיחה בתחומי הקִנים. קשר דומה תיארו Wagner ו-Nicklen [43].
יש להדגיש שנמצאו בני שיח שמתחתם לא היו קִני נמלים, כגון הקזוח התלת-קרני, עובדה המוסברת בריכוז האלקלואידים בצמח שמונע את התבססות הקִנים [17].
השפעתן החיובית של הנמלים על סביבתן ניכרת גם במספר הרב של קִני נמלים בכתמי הנוף המורכבים בהשוואה לכמות המועטה יותר של השיחים בכתמי הנוף המורכבים. כלומר, הנמלים הן "מארחות" טובות יותר לקבוצות ביולוגיות רבות נוספות בהשוואה לבני השיח [26].
אחד הגורמים המגבילים ביותר של תפקוד המערכת האקולוגית המדברית הוא יכולת שימור המים, שחיונית עקב אי-סדירות המשקעים (מבחינת כמותם ופיזורם), ומאפיינת אזורי מדבר [44]. השפעתן של הנמלים על מאפיין זה הודגמה בערכי קיבול שדה דומים בקִנים ובכתמי בני השיח (וגבוהים בכ-5% מאלה של התווך שבין הכתמים), על-ידי הגדלת השטח היחסי שמתפקד כמשמר מים באזורים צחיחים [22]. נוסף על כך, הימצאות חומרי הזנה בריכוז גבוה (אף בהשוואה לכתמי השיח) בשילוב נתוני קיבול השדה מדגישה את חשיבות הקִנים כמִבלעים לשימור מים ולשימור חומרי ההזנה הנאספים מהמקורות מסביב במשך השנה (התאוריה מבלע-מקור [24]) ומפעילות הנמלים בקן [3]. לאחר הגשמים מחלחלים חומרים אלה בדיפוזיה אופקית בתת-הקרקע לכל השטח מסביב [24].
בהסתמך על התאוריה 'מבלע-מקור' ועל התוצאות משטח המחקר לגבי תפרוסת כתמי הנוף השונים המשמשים מבלעים, נמצא שקִני הנמלים הוסיפו כ-55 ק"ג לדונם ביומסה עונתית בהשוואה לביומסה שהייתה מצופה אילו השטח היה מכיל רק תווך (בהתאם לחישוב, היה מצופה שיכיל רק 80 ק"ג לדונם), ולכתמי בני השיח (שתרמו 23 ק"ג לדונם נוסף על הערך המחושב מהתווך בלבד). כמו כן, שטחי הקִנים הוסיפו כ-840 גר' חנקן לדונם לערך המצופה אילו השטח היה מכיל תווך בלבד (בתווך שבין הכתמים נמצאו 31.1 גר') ותוספת של 187.2 גר' פחמן אורגני לדונם (בתווך נמצאו 4.53 ק"ג) [2].
התוצאות המוצגות ממחקר זה התקבלו הן בקִני נמלים פעילים הן בקִנים נטושים. תופעה זו מקבילה לתוצאות מחקר שנערך על השפעתם של כתמי שיח מתים [20], ויכולה לרמוז על השפעתם של קִני נמלת הקציר על הסביבה, אף בטווח הארוך. עם זאת, יש לציין כי מבחני חומרי ההזנה נערכו על קרקע שנאספה באביב. אין ודאות שהתוצאות שהתקבלו מייצגות את הקורה בעונות אחרות, עקב הדינמיות השונה של קִני הנמלים וכתמי השיח [46].
דרך השפעה נוספת על פוריות הסביבה, המיוחסת לשפכי הקרקע מהקן ולסחיפותם הגבוהה [12], נצפתה לאחר הגשמים המרוכזים בינואר 2013 (בין ה-8 ל-10 בינואר ירדו 65 מ"מ גשם, לפי נתונים של בעל החווה), והתבטאה בשפכי אדמה משטחי הקִנים לתווך שבין הכתמים שבמורדותיהם. בבדיקה שבוצעה בנקודה שלא עובדה בוואדי עתיר ביולי 2014, נמצא שבשפכי הקרקע המתקבלים מפעילות הנמלים מצוי חומר אורגני בריכוז 14.4% בהשוואה ל-2.2% בלבד בסביבה הסמוכה הרחוקה משטחי הקִנים (איור 5ד). תוצאות אלה מרחיבות את הידע מהספרות על תרומת הנמלים לפוריות הסביבה [43].

איור 5
השפעתן המרחבית של נמלי הקציר על שטח בתהליך שימור (לאחר שנה), תצפיות משלימות בוואדי עתיר, 2014
א. מבט כללי: מעבר פרטים מהמין נמלת הקציר החולית משטח לא מופר לשטח שעבר עיצוב קרקע נרחב (לימן); ב. צילום תקריב: הקן צעיר בלימן; ג. צילום תקריב: נתיב חורי כניסות נמלים מהשטח השמור ללימן, אורך 3.5 מ'; ד. שפכי קרקע מקן נמלים המצוי בשטח שמור (חווה ביזנטית).
התוצאות שתוארו עד כאן מדגימות את הדינמיקה של קִני הנמלים לאחר כ-20 שנות שימור הקרקע מרעיית יתר. תצפיות משלימות המייצגות את הקורה לשטח מופר בתהליכי שימור, מודגמות באיור 5. באיור מתואר נתיב חורי כניסות נמלים מהשטח הלא מעובד לתוך הלימן שעבר עיבוד קרקע נרחב, שבתוכו התבסס קן נמלים צעיר (יש לציין שמעבר נמלים זה מוגדר כ-patrolling [15], אך לא תואר בין שטחים בעלי ממשק ניהול שונה). תצפיות אלה בוואדי עתיר מחזקות את הממצאים שהתקבלו בשטח המחקר העיקרי בחוות יתיר, לגבי הדינמיקה של הנמלים בשטח שבראשיתו היה מופר (כתוצאה מרעיית יתר) ומצוי בתהליך שימור.
ניתן לסכם את השפעתן של הנמלים על המערכת האקולוגית באזורי מדבר באמצעות הסכֵמה באיור 6.

איור 6
יחסי קִנים-צמחים עונתיים-בני שיח באזורי מדבר
שלב א: נמלים מקן המקור מתיישבות בשטחים השונים בקִנים צעירים (ראו איור 5א–ג, Gordon [15] ו- Schultheissואחרים [33]).
שלב ב: בעוד שהקִנים הצעירים בשטחים החשופים לרעיית יתר כמעט ואינם מתפתחים (ראו איור 1ג), בשטחי הרעייה המתונה יוצרת פעילות הנמלים כתמים עשירים במשאבים ותאחיזת מים גבוהה (ראו איור 4א–ד ו-Beattie and Culver [3]).
שלב ג: חומרי ההזנה ותאחיזת המים הגבוהים בשטחי הקִנים מעודדים את הייצור הראשוני של הביומסה של הצמחים העונתיים (שלב ביניים המשקם את המערכת עד להתנחלות שיחים) (ראו איור 2 ואיור 3).
שלב ג2: Davidson ו-Morton טוענים שחומרי ההזנה המצויים בקִני הנמלים מעודדים ישירות את ההתבססות והגדילה של מספר מיני שיטים באוסטרליה, אף ללא קשר לייצור ביומסה של עשבוניים [9]. טענה זאת, אם תוכח מדעית על מיני בני-שיח ועצים אחרים, יכולה להאיר פן ייחודי ומרתק לגבי השפעת הנמלים על הסביבה.
שלב ד: שלב התנחלות השיחים.
שלב ה: כתוצאה מהתנחלות השיחים והרחבת חופתם פעילות הנמלים בקן ובכתם הנוף מתרחבת (ראו איור 1ב ואיור 1ה) ,עולה פוריות כתם הנוף (ראו איור 1ד), מתרבים השיחים (ראו איור 1ו), נשלחות נמלים לבניית קנים חדשים (ראו איור 1א ו- Mailleux ואחרים [25]) ומתקיימת השפעה על הסביבה בהסתמך על התאוריה מבלע-מקור [24] ובאמצעות שפכי הקרקע (ראו איור 5ד).
מסקנות
מחקר זה התמקד בהשפעת נמלת הקציר החופית והשחורה על מאפייניהם המיקרו- טופוגרפיים של שטחי מדבר, תוך שילוב ניתוחים סטטיסטיים מרחביים ייחודיים. יכולת ההִנדוס הרבה של נמלת הקציר מתבטאת בהגדלת השטח היחסי שמתפקד כמשמר מים, בהעלאת פוריות הקרקע ויצרנותה, וכפועל יוצא מכך – בזירוז השתקמות השטח. עם זאת, תחום המחקר ראשוני, ודרוש מחקר רב נוסף.

תודות
לד"ר ארמין איונסקו מאוניברסיטת תל-אביב על הגדרת מיני הנמלים. לד"ר ברט בוקן מאוניברסיטת בן-גוריון בנגב על הגדרת מיני הצמחים. למשפחות אורן וארן, בעלי חוות יתיר, על העזרה בביצוע המחקר. לעמותת "ואדי עתיר" על ביצוע המחקר בשטחם. המאמר מוקדש לחדווה ויצחק גמינדר על כל התמיכה והעידוד במשך השנים.
- לרעיית יתר יש השפעה שלילית על המערכת האקולוגית בנגב ובאזורי מדבר אחרים בעולם.
- נמלי הקציר כ’מהנדסות סביבה’ עשויות לסייע בשיקום שטחי שיחים בתהליכי התדרדרות.
- התיישבות נמלים יוצרת כתמים עשירים במשאבים, המגדילים את הייצור הראשוני של ביומסת העשבוניים.
- יחסי המשוב בין שיחים לנמלים מגדילים את פוריות המערכת האקולוגית.
- לנמלי הקציר פוטנציאל לתרום לשיקום שטחים מדבריים פגועים.
המערכת
מקורות
- סולר ש, רון מ, פרלמן י ורמון א. 2006. חבל יתיר – סקר, ניתוח והערכה של משאבי טבע, נוף ומורשת האדם. מכון דש"א.
- Aronson J, Floret C, Floc'h E, et al. 1993. Restoration and rehabilitation of degraded ecosystems in arid and semi-arid lands. I. A view from the south. Restoration Ecology 1(1): 8-17.
- Beattie AJ and Culver DC. 1983. The nest chemistry of two seed-dispersing ant species. Oecologia 56(1): 99-103.
- Belsy J. 1992. Effects of grazing, competition, disturbance and fire on species composition and diversity in grassland communities. Journal of Vegetation Science 3(2): 187-200.
- Bernhard-Reversat F. 1982. Biogeochemical cycle of nitrogen in a semi-arid savanna. Oikos 38: 321-332.
- Byers JE, Cuddington K, Jones CG, et al. 2006.Using ecosystem engineers to restore ecological systems. Trends in Ecology and Evolution 21(9): 493-500.
- Cammeraat LH, Willott SJ, Compton SG, and Incoll LD. 2002. The effects of ants' nests on the physical, chemical and hydrological properties of a rangeland soil in semi-arid Spain. Geoderma 105(1-2): 1-20.
- de Bruyn LA. 1999. Ants as bioindicators of soil function in rural environments. Agriculture Ecosystem and Environment 74(1-3): 425-441.
- Davidson DW and Morton SR. 1984. Dispersal adaptations of some Acacia species in the Australian arid zone. Ecology 65(4): 1038-1051.
- Dosta´l P, Br'eznova M, Kozlıckova V, et al. 2005. Ant-induced soil modification and its effect on plant below-ground biomass. Pedobyologia 49: 125-137.
- Eswaran H, Van Den Berg E, and Reich P. 1993. Organic carbon in soils of the world. Soil Science Society of America Journal 57(1): 192-194.
- Farji-Brener AG and Ghermandi L. 2004. Seedling recruitment in a semi-arid Patagonian steppe: Facilitative effects of refuse dumps of leaf-cutting ants. Journal of Vegetation Science 15(6): 823-830.
- Farji-Brener AG. 2005. The effect of abandoned leaf-cutting ant nests on plant assemblage composition in a tropical rainforest of Costa Rica. Ecoscience 12(4): 554-560.
- Goldets C and Boeken B. 2004. Moderate sheep grazing in semiarid shrubland alters small-scale soil surface structure and patch properties. Catena 65: 285-291.
- Gordon DM. 1987. Group-level dynamics in harvester ants: Young colonies and the role of patrolling. Animal Behaviour 35(3): 833-843.
- Greenwood KL and McKenzie BM. 2001. Grazing effects on soil physical properties and the consequences for pastures: A review. Australian Journal of Experimental Agriculture 41(8): 1231-1250.
- Hamada H, Mohammed B, Massiot G, et al. 2004. Alkylated isocoumarins from Pituranthos scoparius. Natural Product Research 18(5): 409-413.
- Holzapfel C and Mahall BE. 1999. Bidirectional facilitation and interference between shrubs and annuals in the Mojave Desert. Ecology 80(5): 1747-1761.
- Jones CG, Lawton JH, and Shachak M. 1994. Organisms as ecosystem engineers. Oikos 69: 373-386.
- Jouquet P, Mathieu J, Choosaï C, and Barot S. 2007. Soil engineers as ecosystem heterogeneity drivers. In: Munoz SI (Ed). Ecology Research Progress. New York: Nova science publishers.
- Lacey JR, Marlow CB, and Lane JR. 1989. Influence of spotted knapweed (Centaurea maculosa) on surface runoff and sediment yield. Weed Technology 3(4): 627-631.
- Le'onard J and Rajot JL. 2001. Influence of termites on runoff and infiltration: Quantification and analysis. Geoderma 104: 17-40.
- Lin Y, Han G, Zhao M, and Chang SX. 2010. Spatial vegetation patterns as early signs of desertification: A case study of a desert steppe in Inner Mongolia, China. Landscape Ecology 25(10): 1519-1527.
- Ludwig JA and Tongway DJ. 1995. Spatial organisation of landscapes and its function in semi-arid woodlands. Australian Landscape Ecology 10(1): 51-63.
- Mailleux AC, Deneubourg JL, and Detrain C. 2003. How does colony growth influence communication in ants? Insectes Sociaux 50(1): 24-31.
- Mor-Mussery A, Leu S, and Budovsky A. 2013. Application of new analytical method for analyzing the areal effects of various biological groups in arid shrublands. Arid Land Research and Management 28(2): 232-241.
- Mor-Mussery A and Leu S. 2012. Project Wadi Attir, 2012 Soil enhancement program, research report 2011/2012.
- Olsvig-Whittaker L, Frankenberg E, Perevolotsky A, and Ungar ED .2006. Grazing, overgrazing and conservation: Changing concepts and practices in the Negev rangelands. Sécheresse 17(1): 195-199.
- Pablo CA and Martin A. 2006. Effects of grazing on patch structure in a semi-arid two-phase vegetation mosaic. Journal of Vegetation Science 16(1): 57-66.
- Rico-Gray V and Oliveira PS. 2007. The ecology and evolution of ant-plant interactions. USA: University of Chicago press.
- Sarah P. 2002. Spatial patterns of soil moisture as affected by shrubs, in different climatic conditions. Environmental Monitoring and Assessment 73: 237-241.
- Sava R. 1994. Guide to sampling, air, water, soil and vegetation for chemical analysis. Environmental Protection Agency, Department of pesticide regulation, Environmental Monitoring and pest management branch. California.
- Schultheiss P, Schwarz S, and Wystrach A. 2010. Nest relocation and colony founding in the Australian desert ant, Melophorus bagoti type Lubbock (Hymenoptera: Formicidae). Psyche: A Journal of Entomology 2010.
- Shachak M and Jones CG. 1995. Ecological flow chains and ecological systems: Concepts for linking species and ecosystem perspectives. In: Jones CG (Ed). Linking Species & Ecosystems. Springer® press US.
- Schiller G and Karschon R. 1974. Microclimate and recreational value of tree plantings in deserts. Landscape Planning 1: 329-337.
- Sidahmed AE. 1996. The rangelands of the arid/semi-arid areas: Challenges and hopes for the 2001 Symposium D: Range Management. The International Conference on Desert Development in the Arab Gulf Countries. KISR, Kuwait 23-26 March 1996.
- Stavi I, Ungar U, Lavee H, and Sarah P. 2009. Livestock modify ground surface microtopography and penetration resistance in a semi-arid shrubland. Arid Land Research and Management 23: 237-247.
- Steinberger Y, Leschnert H, and Shmida A. 1992. Activity pattern of harvester ants (Messor spp.) in the Negev desert ecosystem. Journal of Arid Environment 23: 169-176.
- Swanson J. 2008. Local grazing schemes: Working towards sustainable grazing systems – from pastoralism to wildlands. A best practice guide. 2nd ed. The Grazing Animals Project.
- Tschinkel WR. 2004. The nest architecture of the Florida harvester ant, Pogonomyrmex badius. Journal of Insect Science 4: 21.
- USDA. 1999. Soil taxonomy – A basic system of soil classification for making and interpreting soil surveys ,2nd ed. United State department of agriculture Natural Resources Conservation Service.
- Vonshak M and Lonescu-Hirsch A. 2009. A checklist of the ants in Israel (Hymenoptera: Formicidae). Israel Journal of Entomology 39: 33-57.
- Wagner D and Jones JB. 2006. The impact of harvester ants on decomposition, N mineralization, litter quality, and the availability of N to plants in the Mojave Desert. Soil Biology and Biochemistry 39(8): 2593-2601.
- Ward D, Ngairorue BT, Apollus A, and Tjiveze H. 2000. Perceptions and realities of land degradation in arid Otjimbingwe, Namibia. Journal of Arid Environments 45(4): 337-356.
- Wilby A and Shachak M. 2004. Shrubs, granivores and annual plant community stability in an arid ecosystem. Oikos 106(2): 209-216.
- Zaady E. 2005. Seasonal change and Nitrogen cycling in a patchy Negev desert: A review. Arid Land Research and Management 19: 111-124.
- Zaady E, Yonatan R, Shachak M, and Perevolotsky A. 2001. The effects of grazing on abiotic and biotic parameters in a semiarid ecosystem: A case study from the Northern Negev Desert, Israel. Arid Land Research and Management 15(3): 245-261.



 מהדורה מודפסת
מהדורה מודפסת








