
חזית המחקר
כיצד הפסקת רעיית בקר משפיעה על התאוששות צומח ים תיכוני עשבוני?
חיים קיגל המכון למדעי הצמח והגנטיקה בחקלאות, הפקולטה לחקלאות, מזון וסביבה, האוניברסיטה העברית בירושלים
מרסלו שטרנברג המחלקה לביולוגיה מולקולרית ואקולוגיה של צמחים, הפקולטה למדעי החיים, אוניברסיטת תל–אביב ([email protected])

מבט על חלקות הנתונות לרעייה בצד הצפוני של חוות המחקר כרי דשא בגליל העליון | צילום: קרלי גולודיאץ
מאת
קרלי גולודיאץ
המכון למדעי הצמח והגנטיקה בחקלאות, הפקולטה לחקלאות, מזון וסביבה, האוניברסיטה העברית בירושלים; המחלקה לביולוגיה מולקולרית ואקולוגיה של צמחים, הפקולטה למדעי החיים, אוניברסיטת תל-אביבחיים קיגל
המכון למדעי הצמח והגנטיקה בחקלאות, הפקולטה לחקלאות, מזון וסביבה, האוניברסיטה העברית בירושליםמרסלו שטרנברג
המחלקה לביולוגיה מולקולרית ואקולוגיה של צמחים, הפקולטה למדעי החיים, אוניברסיטת תל–אביבמאת
קרלי גולודיאץ
המכון למדעי הצמח והגנטיקה בחקלאות, הפקולטה לחקלאות, מזון וסביבה, האוניברסיטה העברית בירושלים; המחלקה לביולוגיה מולקולרית ואקולוגיה של צמחים, הפקולטה למדעי החיים, אוניברסיטת תל-אביבחיים קיגל
המכון למדעי הצמח והגנטיקה בחקלאות, הפקולטה לחקלאות, מזון וסביבה, האוניברסיטה העברית בירושליםמרסלו שטרנברג
המחלקה לביולוגיה מולקולרית ואקולוגיה של צמחים, הפקולטה למדעי החיים, אוניברסיטת תל–אביברעייה היא חלק בלתי נפרד מהנוף הים תיכוני זה אלפי שנים. בתגובה לשינויים בתנאי הרעייה במשך השנים, התחוללו שינויים במבנה הצומח ובתפקודו, מהרכב המינים ועד לתהליכים מרכזיים במערכת האקולוגית. במחקר הנוכחי נבחנה תגובת הצומח העשבוני הים תיכוני לירידה בלחץ הרעייה ולהפסקת הרעייה בטווח זמן קצר וארוך (3 שנים וכ-30 שנים). המחקר בוצע בין השנים 2003–2005 בחוות המחקר כרי דשא בגליל העליון, שבה עדר בקר לבשר. בחלקות הנתונות לרעייה נבנו גדורות למניעת הרעייה כדי לבחון את התאוששות הצומח בטווח הקצר. לחיזוי השינויים בחברת הצומח ובתפקוד המערכת האקולוגית נבדקו במחקר שינויים בהרכב הצומח, תפקודי המערכת האקולוגית (ייצור ביומסה ופירוק הנשר הצמחי), ושימוש בתכונות LHS של צמחים (L=השטח הסגולי של העלה [SLA], H=גובה הצמח, S=מסת הזרע). נמצא כי בטווח הקצר גרמה הפסקת הרעייה להתאוששות מהירה של הדגניים הרב-שנתיים הגבוהים ולהשתלטותם על השטח תוך שלוש שנים. נוצר הרכב צומח דומה לזה של החלקות שהיו ללא רעייה תקופה ממושכת, ונרשמה עלייה ביצרנות, אולם ללא השפעה על קצב פירוק הנשר הצמחי. מבחינת תכונות ה-LHS, עלה חלקם של מינים גבוהים, בעלי זרעים גדולים, אך עם SLA קטן יותר. תכונות ה-LHS הציגו היטב את השינויים בצומח בעקבות הפסקת הרעייה, והצליחו לחזות את השינויים בייצור הביומסה. ניתן להסיק כי הצומח העשבוני הים תיכוני מתאפיין בתחלופה במינים הדומיננטיים בתגובה לשינויים ברעייה. הדינמיקה של המינים הדומיננטיים היא מדד טוב לשינויים בהרכב הצומח הכללי, לייצור הביומסה ולתכונות ה-LHS. מסקנה נוספת היא שהצומח מתאפיין בכושר התאוששות מהיר אחרי הפסקת הרעייה, המצביע על יציבות גבוהה של המערכת בפני רעיית בקר, זאת כנראה בעקבות ההיסטוריה ארוכת השנים של הרעייה באזור כולו.
על קצה המזלג
- רעייה של צאן ובקר, שהיא חלק מהנוף הים תיכוני כבר אלפי שנים, משפיעה על מבנה הצומח ועל הרכבו.
- נבחנה תגובת צומח עשבוני בשטח מרעה לצמצום הרעייה ואף להפסקתה, בטווח זמן קצר (3 שנים) וארוך (30 שנים).
- הניסוי נערך בחוות המחקר כרי דשא בגליל העליון, הנמצאת תחת משטר רעייה מבוקר של עדר פרות.
- נמצא כי תוך זמן קצר (3 שנים) מהפסקת הרעייה התאושש הצומח וחזר למצבו כפי שהיה ללא רעייה.
- ההתאוששות המהירה מצביעה על יציבות המערכת האקולוגית בפני רעייה, כנראה בעקבות ההיסטוריה הארוכה של הרעייה באזור כולו.
המערכת
הלכה למעשה
מבוא
רעייה של בעלי החיים מבויתים היא חלק בלתי נפרד מהנוף הים תיכוני זה אלפי שנים [15]. השינויים שחלו במשך השנים במשטרי הרעייה, ביחד עם תנודות האקלים באזור, השפיעו על מבנה הצומח ועל הרכבו, ומתבטאים ברמות ארגון שונות של חברת הצמחים [15]. לדוגמה, שינויים במשטר רעיית בקר במערכת העשבונית של צפון הגליל והגולן משפיעים על הרכב הצומח ועל תהליכים מרכזיים במערכת האקולוגית, כגון ייצור ביומסה (יצרנות) ופירוק נשר (חומר צמחי יבש) הדרוש למִחזור חומרים מְזינים (nutrients) במערכת [5].
ניתן לאפיין את הצמחייה העשבונית לפי קבוצות תפקודיות (קבוצות מינים בעלי תכונות דומות) [18, 19]. שינויים במשטר הרעייה יכולים לשנות את הרכב הצומח היות שלמינים ולקבוצות התפקודיות יש אסטרטגיות שונות להתמודדות עם הרעייה ועם השינויים שהיא גורמת בתנאי התאורה ובמשאבי הקרקע [15]. נוסף על כך, הרעייה היא סלקטיבית – בעלי החיים מעדיפים מינים מסוימים ונמנעים מאכילת מינים אחרים. משום כך למשל, ירידה בלחץ רעיית בקר או הפסקת הרעייה מגדילות את התחרות על אור במערכת העשבונית בגליל, וזאת מאחר שנוצרת עלייה בכיסוי הדגניים הגבוהים [15] המועדפים על-ידי הפרות, והם דוחקים קבוצות אחרות, כגון דגניים נמוכים חד-שנתיים [18]. עלייה בכיסוי הדגניים הגבוהים יכולה להוביל לעלייה בייצור הביומסה, אך גם להאטה בפירוק הנשר כתוצאה מריבוי רקמות תמיכה המכילות ליגנין (רקמות חיזוק קשיחות) במינים הגבוהים [13]. לעומת זאת, קצב פירוק הנשר הנוצר מצומח תחת רעייה עשוי להיות מהיר יותר בגלל הקטנת חלקן של רקמות תמיכה במינים הנמוכים השכיחים יותר ברעייה [15, 18], ועקב תכולה גבוהה של חלבונים באיברים הצעירים שמכילים פחות ליגנין, ושמתפתחים בצמחים שנפגעים מרעייה [8].
גישה אחרת לבחינת השינויים בצומח נעזרת בתכונות תפקודיות, כלומר בתכונות הקשורות לתפקודים מסוימים של הצמח. לדוגמה, השטח הסגולי של העלה (SLA – יחידת מסה ליחידת שטח) קשור לקצב הגידול של הצמח, גובה הצמח מייצג את יכולת התחרות שלו, ומסת הזרע משפיעה על הנביטה ועל התבססות הנבטים. עקב קשרים אלה, הוצע להתייחס לכל אחת מהתכונות התפקודיות הללו כמייצגת ממד בלתי תלוי של שונות בחברת הצמחים – סכֵמת (Leef-Height-Seed) LHS [21]. במקרה של ירידה בלחץ הרעייה או הפסקת הרעייה, הגורמות לעלייה בצפיפות הצמחים, צפוי כי: א) הצמח יגבה כתוצאה מתחרות על אור עם צמחים שכנים; ב) מסת הזרע תעלה תוך הגדלת הסיכוי להצצה ולהתבססות הנבטים בתנאים של צל והצטברות נשר צמחי; ג) SLA יקטן עקב השקעה מוגברת ברקמות תמיכה בעלה [22]. מחקרים שבחנו סכֵמה תאורטית זו מצאו קשר בין התכונות לבין התפקודים האלה ברמת המין [2, 10, 14]. במחקרנו, בחנו את יישום סכֵמת ה-LHS גם ברמת החברה [14], על-ידי חישוב ערכים משוקללים של תכונות הצמחים בחברה, לפי תרומתם היחסית של המינים הנפוצים לייצור הביומסה הכללית בחברת הצמחים [4]. גישה זו מבוססת על השערת יחסי הביומסה (biomass ratio hypothesis) [7], המניחה שהמינים הנפוצים הם בעלי השפעה גבוהה על תהליכים ברמת המערכת האקולוגית. כל שינוי בתרומה היחסית של המינים הללו לביומסה ישנה את הממוצע המשוקלל, וכך ניתן יהיה לבחון את תגובת הצומח לשינויים בתנאי הרעייה. בדקנו גישה זו מאחר שבמערכת הנחקרת משפיעים שינויים ברעייה בעיקר על השפע היחסי של המינים השונים, ולא על תחלופה של מינים [18].
מאחר שתכונות תפקודיות מייצגות תפקודים שונים של הצמחים הנפוצים במערכת האקולוגית [21, 22], השאלה המתבקשת היא: האם ניתן להשתמש בתכונות תפקודיות של צמחים לחיזוי שינויים בתהליכים ברמת המערכת האקולוגית בעקבות שינויים בתנאי הרעייה [4, 12]? אם כן, נקבל הוכחה שהשערת יחסי הביומסה תקפה במציאות, ונוכל להסתפק במדידת מספר תכונות תפקודיות במיני הצמחים הנפוצים, במקום בדגימה אינטנסיבית של כלל הביומסה והנשר בצומח.
במחקרנו בחנו את התגובות של המערכת העשבונית הים תיכונית בגליל העליון לתנאים שונים של רעיית בקר, במטרה להבין את יחסי הגומלין בין תכונות מיני הצמחים, הרכב הצומח והתהליכים ברמת המערכת האקולוגית.
שיטות
תיאור האתר ומערך הניסוי
המחקר בוצע בחוות כרי דשא בגליל העליון (570 מ"מ גשם בממוצע). הצומח הוא עשבוני ים תיכוני, לרוב חד-שנתי, עם שכיחות גבוהה של דגניים. המין הבולט הוא שעורת הבולבוסין (Hordeum bulbosum), דגן רב-שנתי גבוה. עונת הצמיחה מתחילה עם הגשם הראשון (אוקטובר–נובמבר) ונמשכת עד אפריל–מאי. הצומח בכרי דשא נמצא תחת משטר רעייה מבוקר של עדר בקר לבשר [18]. לצורך המחקר בחרנו שני לחצי רעייה – לחץ גבוה (CH; 9 דונם לפרה) ולחץ מתון (CM; 18 דונם לפרה) – וקבענו לכל לחץ רעייה שתי חלקות. נוסף על כך, בחרנו שתי חלקות המוגנות מרעייה כ-30 שנה ויותר לפני תחילת הניסוי (LP). בכל אחת מהחלקות הנתונות לרעייה נבנו ב-2003 חמש גדורות של 10X10 מטר כדי לבחון את התאוששות הצומח בטווח הקצר בהיעדר רעייה.
דגימה ומדידות
במשך שלוש שנים (2003–2005) דגמנו את הצומח העשבוני בתוך הגדורות ובחלקות המוגנות מרעייה. נלקחו חמש דגימות של 25X25 ס"מ בשיא עונת גידול הצומח (אפריל). במעבדה עברו הדגימות מיון [1], ייבוש ( 70°C במשך 48 שעות), שקילה לפי מין וקביעת ביומסה כללית. בקיץ 2003 אספנו נשר במרכז כל גדורה ובחלקות המוגנות לצורך ניסוי פירוק הנשר. בכל חלקה ערבבנו את הנשר מכל הגדורות. במעבדה הכנו שקיות רשת (נקבים ברוחב 1.1 מ"מ) בגודל 15X15 ס"מ והכנסנו לכל שקית 4 גרם נשר. 15 שקיות הונחו בתוך כל הגדורות ובחלקות המוגנות לפני תחילת עונת הגשמים השנייה (אוקטובר 2003). במשך שלוש שנים, בקיץ, הוצאו 5 שקיות מכל גדורה לשקילה במעבדה. קצב פירוק הנשר נקבע על-ידי איבוד מסת הנשר מהשקית. בתחילת הניסוי עברו 5 דגימות של נשר מכל חלקה אנליזה כימית לקביעת ההרכב הכימי של הנשר בטיפולים השונים. לפני תחילת העונה השלישית של דגימת הביומסה הירוקה (אוקטובר 2004) נדגם הנשר הצמחי לפי השיטה של דגימת הביומסה הירוקה, ונדגמו 5 דגימות מחוץ לכל אחת מהגדורות, וזאת נוסף על הדגימה בתוך הגדורות.
ב-2003 וב-2004 נדגמו תכונות ה-LHS בשיא עונת גידול הצומח, במינים העיקריים שתרמו ל-80% מכלל הביומסה בכל טיפול באותן שנים – 15 מינים. לצורך מדידת ה-SLA וגובה הצמח נבחרו צמחים באקראי בתוך הגדורות ובחלקות המוגנות – בכל טיפול 6–10 צמחים לכל אחד מ-15 המינים. מכל צמח נבחר העלה המפותח הצעיר ביותר לצורך חישוב ה-SLA; שטח העלה נמדד, העלה יובש, ולאחר מכן נשקל. גובה הצמח נקבע על-ידי מדידת הצמח מבסיס הגבעול כולל תפרחות. מסת הזרע נמדדה מזרעים שנאספו מדגימות הביומסה, ומזרעים שנאספו בחלקות בקיץ, משום שהמינים הקוצניים מבשילים מאוחר יותר. לכל מין נלקחו 6–10 דגימות של עשרה זרעים, ומסה של זרע אחד חושבה מהמסה הממוצעת של אותם עשרה זרעים. עבור כל אחת משלוש התכונות חושבו ערכים משוקללים לכל טיפול, לפי תרומתו של כל אחד מ-15 המינים הנפוצים לביומסה בטיפולים שונים.
ניתוחים סטטיסטיים
בחנו את השפעת טיפולי הרעייה על כל המדדים באמצעות ניתוח שונות (ANOVA). נתוני ייצור הביומסה ואיבוד המסה בניסוי פירוק הנשר נותחו עם ניתוח שונות בתכונת מדידות נשנות (repeated measures) עם שנה כחזרה, וכללו את יחסי הגומלין בין טיפול ושנה. ניתוחי השונות נבנו כמודלים מקננים (nested models), והרמה הנמוכה ביותר שימשה כשגיאה ניסיונית. הנתונים עברו טרנספורמציה לפי הצורך (arcsin עבור אחוזים, ln עבור משקל הביומסה והנשר, ותכונות ה-LHS). השתמשנו במתאם מסוג פירסון עבור תכונות ה-LHS, ומסוג ספירמן עבור הערכים המשוקללים של ה-LHS והמתאמים ביניהם לבין לחץ הרעייה. כל ניתוחי השונות והמתאמים בוצעו בתכנת JMP IN 5.1.
ביצענו אורדינציה (שיטה מתמטית לצמצום השונות בנתונים למספר מצומצם של צירים בלתי תלויים) כדי לבחון באיזו מידה הערכים המשוקללים של ה-LHS מסבירים את השונות בין החלקות השונות. כדי לאפשר גם בחינה של הקשר בין ה-LHS ולחץ הרעייה, בחרנו באורדינציה מסוג RDA – Redundancy Analysis, המאפשרת הוספת גורמי סביבה כמשתנים מסבירים. לצורך כך, כללנו את לחץ הרעייה כמשתנה מסביר. האורדינציה בוצעה ב-CANOCO for Windows 4.5, וכללה מבחן תמורה (permutation test) לבחינת הקשר בין לחץ הרעייה והציר הראשון של האורדינציה [20]. הניתוח סיפק גם מתאמים בין ה-LHS וצירי האורדינציה. נתוני הביומסה של 15 המינים נוספו לגרף לצורך פענוח התוצאות. אורדינציה דומה בוצעה כדי לבחון באיזו רמה ייצור הביומסה ופירוק הנשר מסבירים את השונות בין החלקות, כאשר הערכים המשוקללים של ה-LHS שימשו משתנים מסבירים. נתוני היחס בין ליגנין לחנקן נוספו לצורך פענוח התוצאות.
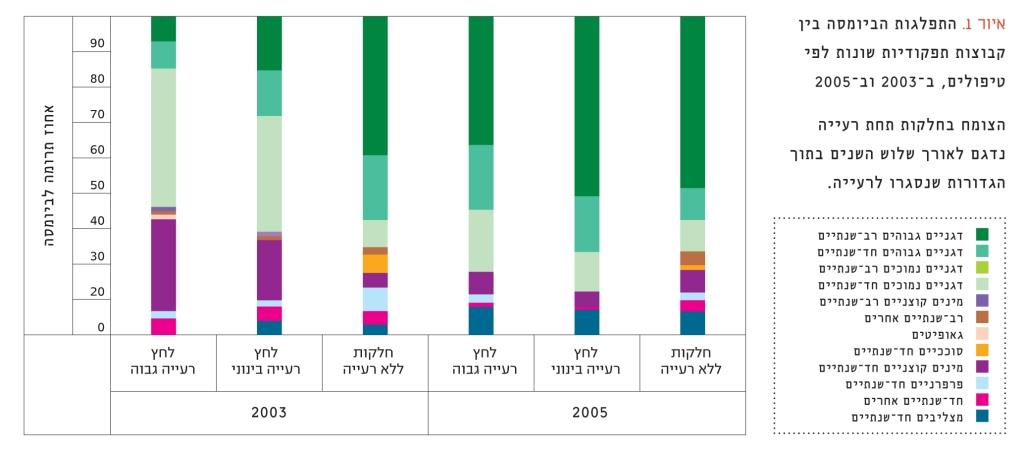
איור 1
התפלגות הביומסה בין קבוצות תפקודיות שונות לפי טיפולים, ב-2003 וב-2005
הצומח בחלקות תחת רעייה נדגם לאורך שלוש השנים בתוך הגדורות שנסגרו לרעייה.
תוצאות
הרכב מינים וביומסה לפי קבוצות תפקודיות
לאחר הפסקת הרעייה בגדורות שנבנו בחלקות עם לחץ הרעייה המתון (CM) והגבוה (CH), התרחשו מספר שינויים בהרכב הצומח (איור 1). עם הזמן חלה ירידה בביומסה היחסית של מינים קוצניים חד-שנתיים ושל דגניים חד-שנתיים נמוכים, והיא הגיעה לרמה שנרשמה בחלקות המוגנות מרעייה לטווח הארוך (LP). לעומת זאת, חלה עלייה בביומסה היחסית של דגניים רב-שנתיים גבוהים (בעיקר שעורת הבולבוסין), וגם היא הגיעה לרמה שנרשמה בחלקות המוגנות. הרכב המינים בחלקות המוגנות לא השתנה באופן משמעותי בין השנים 2003–2005.
ייצור ביומסה כללית ונשר צמחי
ייצור הביומסה ב-2003, שנת הקמת הגדורות, היה גבוה יותר בחלקות המוגנות מרעייה לטווח הארוך מאשר בחלקות הנתונות לרעייה (איור 2א). עד 2005 עלה ייצור הביומסה בגדורות (CM, CH) לרמה שנרשמה בחלקות המוגנות. כמו כן, ייצור הנשר הצמחי היה גבוה יותר במדידות שנערכו בחלקות המוגנות מאשר בגדורות, הן בתוך הגדורות הן מחוץ להן (איור 2ב). באופן כללי, ייצור הנשר בתוך הגדורות היה גבוה מייצור הנשר מחוץ לגדורות.
איכות הנשר הצמחי ופירוקו
הנשר הצמחי בגדורות היה איכותי יותר מאשר בחלקות המוגנות, היות שכמות הליגנין בחלקות המוגנות (7.30%) הייתה גבוהה יותר מאשר בגדורות שהיה בהן לחץ רעייה מתון (4.75%) וגבוה (5.63%); כמו כן, היחס בין ליגנין לחנקן היה גבוה בחלקות המוגנות (11.62) לעומת החלקות שהיה בהן לחץ רעייה מתון (8.17) וגבוה (8.46), אף על פי שלא היה שינוי ברמת החנקן בנשר בין הטיפולים השונים (ממוצע חנקן 0.63%). למרות השינוי באיכות הנשר, לא נמצא הבדל בקצב פירוק הנשר בין הטיפולים השונים לאורך כל שלוש שנות הניסוי (איור 2ג).
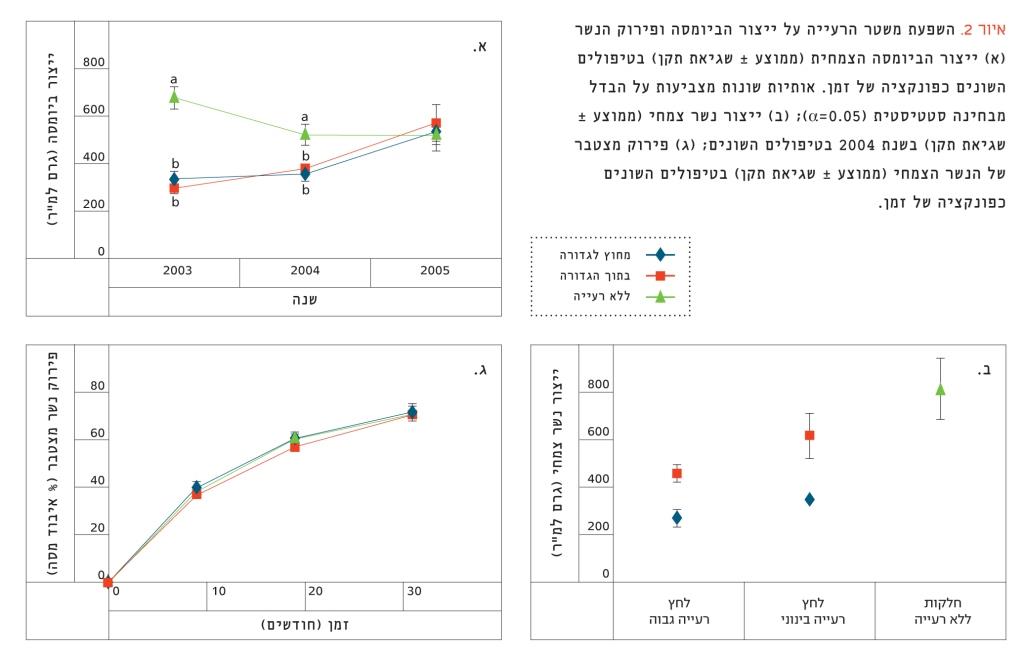
איור 2
השפעת משטר הרעייה על ייצור הביומסה ופירוק הנשר (א) ייצור הביומסה הצמחית (ממוצע ± שגיאת תקן) בטיפולים השונים כפונקציה של זמן.
אותיות שונות מצביעות על הבדל מבחינה סטטיסטית (0.05=α); (ב) ייצור נשר צמחי (ממוצע ± שגיאת תקן) בשנת 2004 בטיפולים השונים; (ג) פירוק מצטבר של הנשר הצמחי (ממוצע ± שגיאת תקן) בטיפולים השונים כפונקציה של זמן.
תכונות תפקודיות של הצמחים
ברמת המין, כל התכונות הגיבו באופן חיובי להפסקת הרעייה (איור 3). תוצאה זו הייתה מובהקת מבחינה סטטיסטית עבור SLA (איור 3א) ועבור מסת הזרע (איור 3ג). לעומת זאת, הערכים המשוקללים הראו ש-SLA יורד עם הפסקת הרעייה ואילו גובה הצמח ומסת הזרע עולים באופן מובהק (איור 3). ברמת המין נמצא מתאם שלילי מובהק בין SLA ומסת הזרע (r=–0.572, P<0.05), ומתאם חיובי מובהק (P<0.05) בין גובה הצמח ומסת הזרע (r=0.517). המתאם השלילי בין גובה הצמח ו-SLA לא היה מובהק (r=–0.346). ברמת החברה נרשמו מתאמים גבוהים בין כל שלוש התכונות. גובה הצמח ומסת הזרע היו במתאם חיובי (r=1.00) וכל אחת הייתה במתאם שלילי עם SLA (r=–0.94 ו-r=–0.93 בהתאמה, עבור גובה הצמח ומסת הזרע). בהתאם לתוצאות ברמת החברה, SLA נמצא במתאם חיובי עם לחץ הרעייה (r=0.837) ואילו גובה הצמח ומסת הזרע הראו מגמה הפוכה (r=–0.956 עבור שתי התכונות). הציר הראשון של האורדינציה הסביר 79% מהשונות וייצג את לחץ הרעייה (איור 4). לכן, מבחני ההתאמה עם תכונות ה-LHS היו מובהקות (r= –0.923, r= 0.829 ו-r=–0.911 עבור SLA, גובה הצמח ומסת הזרע, בהתאמה). הציר השני של האורדינציה הסביר 20% מהשונות. ניתן לראות באיור 4 כי שלוש התכונות יוצרות ציר אחד ברור, שבו גובה הצמח ומסת הזרע נמצאים בקצה האחד, וקשורים לחלקות המוגנות ולמינים הנפוצים בטיפול זה (כגון שעורת הבולבוסין), ואילו SLA נמצא בקצה השני, ומייצג חלקות עם רעייה חזקה, ואת המינים הנפוצים שם.
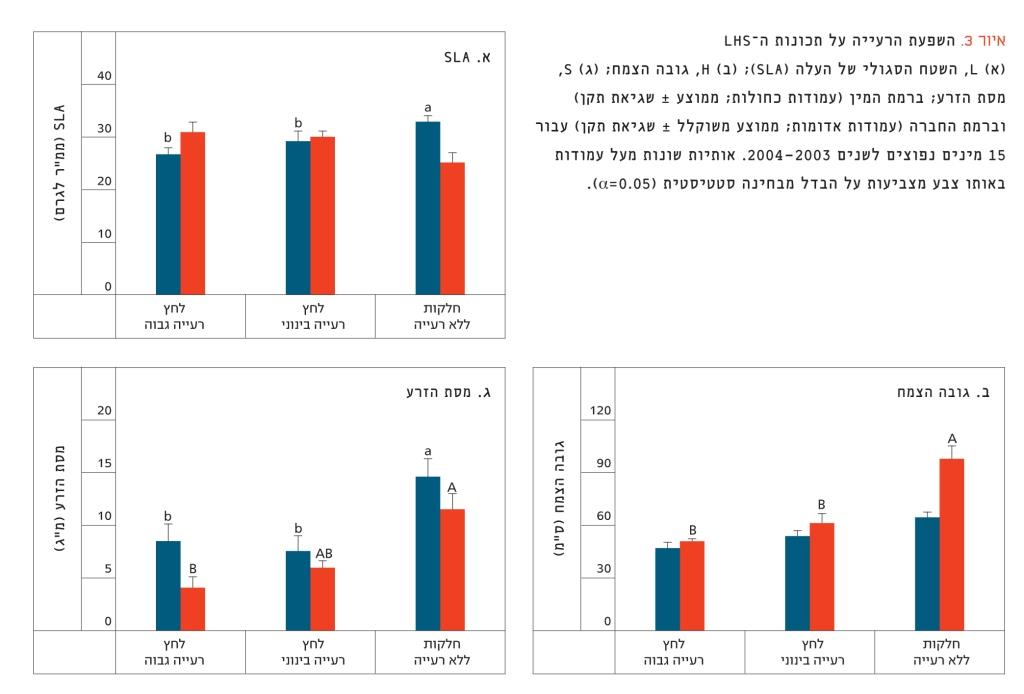
איור 3
השפעת הרעייה על תכונות ה-LHS
(א) L, השטח הסגולי של העלה (SLA); (ב) H, גובה הצמח; (ג) S, מסת הזרע; ברמת המין (עמודות כחולות; ממוצע ± שגיאת תקן) וברמת החברה (עמודות אדומות; ממוצע משוקלל ± שגיאת תקן) עבור 15 מינים נפוצים לשנים 2003–2004. אותיות שונות מעל עמודות באותו צבע מצביעות על הבדל מבחינה סטטיסטית (0.05=α).
הקשר בין תכונות תפקודיות של צמחים ותהליכים ברמת המערכת האקולוגית
איור 5 מציג את הקשר שבין תכונות ה-LHS והתהליכים ברמת המערכת האקולוגית כפי שנמדדו בשנה הראשונה (2003) – ייצור הביומסה וקצב פירוק הנשר. נמצא כי ייצור הביומסה קשור באופן הדוק לתכונות ה-LHS. ייצור ביומסה גבוה בחלקות ללא רעייה מיוחס לצמחים גבוהים עם זרעים גדולים, ואילו ייצור ביומסה נמוך בחלקות עם רעייה מיוחס לצמחים עם SLA גבוה. לעומת התגובה הברורה של ייצור הביומסה, פירוק הנשר לא השתנה בטיפולי הרעייה השונים ולא נמצא קשר בינו לבין אף אחת מתכונות ה-LHS.
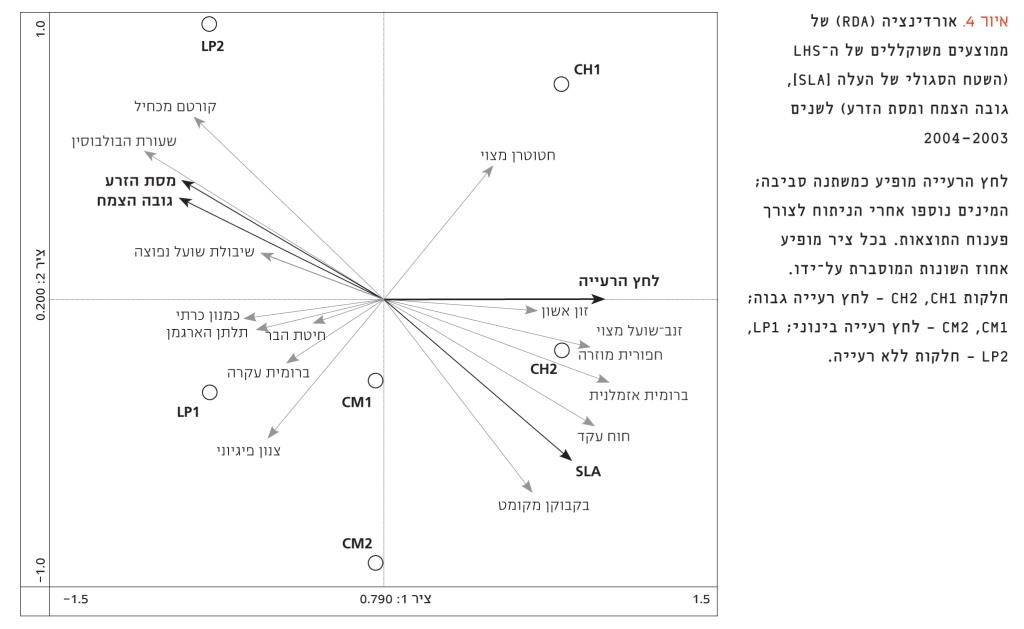
איור 4
אורדינציה (RDA) של ממוצעים משוקללים של ה-LHS (השטח הסגולי של העלה [SLA], גובה הצמח ומסת הזרע) לשנים 2003–2004
לחץ הרעייה מופיע כמשתנה סביבה; המינים נוספו אחרי הניתוח לצורך פענוח התוצאות. בכל ציר מופיע אחוז השונות המוסברת על-ידו. חלקות CH2 ,CH1 – לחץ רעייה גבוה; CM2 ,CM1 – לחץ רעייה בינוני; LP2 ,LP1 – חלקות ללא רעייה.
דיון
מממצאי מחקר זה מצטיירת תמונה כלל-מערכתית של תגובת הצומח העשבוני הים תיכוני בכרי דשא להפסקת הרעייה. תוך שלוש שנים מהשחרור מרעייה (בניית הגדורות) התאוששו הדגניים הגבוהים [18], וגרמו לדיכוי של הדגניים הנמוכים החד-שנתיים והמינים הקוצניים החד-שנתיים. עלייתם של הדגניים הגבוהים בסולם הדומיננטיות גרמה לעלייה ניכרת בייצור הביומסה הכללית של הצומח העשבוני, עד לרמת ייצור המשתווית לזו שנמצאה בחלקות המוגנות מרעייה לטווח הארוך (יותר מ-30 שנה). בד בבד עם העלייה בייצור הביומסה נמצאה עלייה בייצור ובהצטברות של הנשר הצמחי.
השינויים שמתרחשים בהרכב הצומח בעקבות הפסקת הרעייה גורמים לשינויים ברורים בערכים המשוקללים של שלוש תכונות תפקודיות חשובות – השטח הסגולי של העלה (SLA), גובה הצמח ומסת הזרע. העלייה בכיסוי של הדגניים הגבוהים סוגרת את החופה מעל שאר המינים ויוצרת תנאים של תחרות לאור [15]. תנאים אלה גורמים לעלייה בשכיחות המינים הגבוהים ובגובה הכללי של הצומח, ויוצרים עדיפות למינים בעלי זרעים גדולים (לרוב מינים גבוהים), המסוגלים לנבוט דרך שכבה עבה של נשר צמחי ולהתבסס בתנאי צל [19]. כמו כן, כדי להתחרות ולהגיע לגובה רב, הצמחים נדרשים להשקיע משאבים רבים יותר ברקמות תמיכה [3]. התוצאה הסופית היא עלייה בגובה הצמח ובמסת הזרע וירידה ב-SLA. מכיוון שהשינויים בהרכב המינים נובעים משינויים בדומיננטיות בין המינים הנפוצים ולא מתחלופה של מינים בין הטיפולים השונים, השינויים בתכונות התפקודיות ברמת החברה משקפים בצורה הברורה ביותר את הדינמיקה בין המינים בעקבות הפסקת הרעייה [6]. מתוך כך ניתן להסביר מדוע נכשלה סכֵמת ה-LHS ברמת המין. כאשר אין תחלופה של המינים הנפוצים בין הטיפולים, אין הרבה משמעות לממוצע האריתמטי של התכונה. ייתכנו שינויים תוך-מיניים בין הטיפולים ב-SLA ובגובה הצמח [11], אולם שינוי במין שכיח פחות שקול לשינוי במין שכיח יותר, ועלולות להתקבל תוצאות סותרות ולא-הגיוניות, שלא ישקפו את הדינמיקה בין המינים.
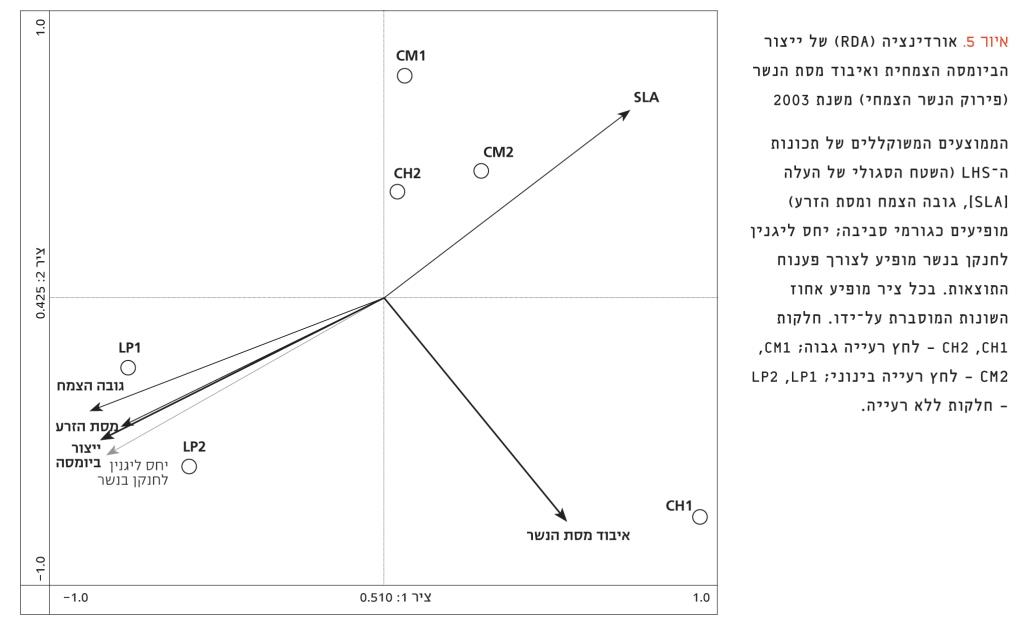
איור 5
אורדינציה (RDA) של ייצור הביומסה הצמחית ואיבוד מסת הנשר (פירוק הנשר הצמחי) משנת 2003
הממוצעים המשוקללים של תכונות ה-LHS (השטח הסגולי של העלה [SLA], גובה הצמח ומסת הזרע) מופיעים כגורמי סביבה; יחס ליגנין לחנקן בנשר מופיע לצורך פענוח התוצאות. בכל ציר מופיע אחוז השונות המוסברת על-ידו. חלקות CH2 ,CH1 – לחץ רעייה גבוה; CM2 ,CM1 – לחץ רעייה בינוני; LP2 ,LP1 – חלקות ללא רעייה.
כפי שהפסקת הרעייה השפיעה באופן ברור ומובהק על ייצור הביומסה, כך ציפינו לראות שינוי בקצב פירוק הנשר הצמחי. הנחנו כי ההבדלים בהרכב המינים בטווח הארוך ייצרו נשר איכותי יותר בחלקות הרעייה לעומת הנשר בחלקות ללא רעייה, בהתבסס על שני תהליכים שונים: ייצור מוגבר של רקמות מבניות ורקמות תמיכה בחלקות המוגנות, המקשות על פירוק הנשר מחד גיסא, ומאידך גיסא, שכיחות גבוהה של מינים נמוכים בעלי קצב גידול מהיר (SLA גבוה) [15] והתחדשות של צמחים אכולים [8] בחלקות הרעייה, שכולם עתירי חומרים מְזינים וקלים לפירוק. אמנם נמצא הבדל סטטיסטי מובהק בתכולת הליגנין (חומר המעכב את פירוק הנשר), שהייתה גבוהה יותר בנשר בחלקות המוגנות, ונמצא הבדל ביחס בין ליגנין לחנקן המשמש מדד נפוץ לאיכות הנשר [17], אולם לא נמצא הבדל בתכולת החנקן. בניגוד לציפיות, תנאי הרעייה לא השפיעו על קצב פירוק הנשר הצמחי. לכך יכולות להיות שתי סיבות אפשריות: כקבוצת-על, הדגניים במערכת עשבונית זו יוצרים 80%–60 מכלל הביומסה, וההרכב הכימי של הנשר אינו משתנה באופן משמעותי בין דגניים נמוכים, גבוהים, חד-שנתיים ורב-שנתיים [9]. נוסף על כך, רמת הליגנין בדגניים ככלל נמוכה מאוד, והשפעתו על קצב פירוק הנשר זניחה. בעקבות חוסר התגובה של קצב פירוק הנשר לשינויים בתנאי הרעייה, לא ניתן להשתמש בתכונות תפקודיות של צמחים כדי לחזות שינויים בקצב פירוק הנשר במערכת עשבונית זו, שהדגניים שולטים בה.
סיכום
חברת הצומח העשבונית מתאפיינת בשינויים בדומיננטיות המינים בתגובה לשינויים בתנאי הרעייה. כתוצאה מכך, הדינמיקה של המינים הדומיננטיים היא מדד טוב לשינויים בחברה כולה, מבחינת הרכב המינים, ייצור הביומסה, התכונות התפקודיות של הצמחים והקשרים בין היבטים אלה. מאפיין חשוב נוסף הוא ההתאוששות המהירה והמרשימה של הצומח אחרי הפסקת הרעייה. שני המאפיינים הללו יחד מצביעים על יציבות גבוהה של המערכת בפני רעיית בקר, הנובעת מההיסטוריה ארוכת השנים של הרעייה באזור כולו [16, 18], ומאפשרים קיום משטר רעייה בר-קיימא ללא פגיעה בתפקוד של מערכת עשבונית זו. נוסף על כך, קיום של לחצי רעייה שונים לצד חלקות ללא רעייה יוצר מעין פסיפס של בתי גידול בנוף, המאפשר שמירה על עושר מינים ועל מגוון מינים גבוה באזור כולו [16].
תודות
מחקר זה הוא חלק מעבודת הדוקטור של קרלי גולודיאץ. ברצוננו להודות לאבי פרבולוצקי ולעמנואל נוי-מאיר (ז"ל) על הייעוץ המקצועי, לאירית קונסנס וליובל ספיר על עזרה בהגדרת הצמחים, להילרי פוט ולסנדרה לבורל על הייעוץ הסטטיסטי, לזלמן הנקין ולצוות בכרי דשא, ולסטודנטים הרבים שעזרו במעבדה ובשדה.
- רעייה של צאן ובקר, שהיא חלק מהנוף הים תיכוני כבר אלפי שנים, משפיעה על מבנה הצומח ועל הרכבו.
- נבחנה תגובת צומח עשבוני בשטח מרעה לצמצום הרעייה ואף להפסקתה, בטווח זמן קצר (3 שנים) וארוך (30 שנים).
- הניסוי נערך בחוות המחקר כרי דשא בגליל העליון, הנמצאת תחת משטר רעייה מבוקר של עדר פרות.
- נמצא כי תוך זמן קצר (3 שנים) מהפסקת הרעייה התאושש הצומח וחזר למצבו כפי שהיה ללא רעייה.
- ההתאוששות המהירה מצביעה על יציבות המערכת האקולוגית בפני רעייה, כנראה בעקבות ההיסטוריה הארוכה של הרעייה באזור כולו.
המערכת
מקורות
- פיינברון–דותן נ ודנין א. 1998. המגדיר לצמחי בר בארץ ישראל. ירושלים: כרטא.
- Garnier E, Bellman A, Navas ML, et al. 2004. The leaf-height-seed plant ecology strategy scheme as applied to species from a Mediterranean old-field succession. In: Arianoutsou and Papanastasis (Eds). Proceedings 10th MEDECOS Conference; 2004 Apr 25-May 1; Rhodes Island, Greece. Rotterdam, NL: Millpress.
- Garnier E, Cordonnier P, Guillerm JL and Sonié L. 1997. Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in Mediterranean old-fields. Oecologia 111: 490-498.
- Garnier E, Lavorel S, Ansquer P, et al. 2007. Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: A standardized methodology and lessons from an application to 11 European sites. Annals of Botany 99: 967-985.
- Golodets C, Kigel J, and Sternberg M. 2010. Recovery of plant species composition and ecosystem function after cessation of grazing in a Mediterranean grassland. Plant and Soil 329: 365-378.
- Golodets C, Sternberg M, and Kigel J. 2009. A community-level test of the leaf-height-seed ecology strategy scheme in relation to grazing conditions. Journal of Vegetation Science 20: 392-402.
- Grime JP. 1998. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. Journal of Ecology 86: 902-910.
- Henkin Z, Landau SY, Ungar ED, et al. 2007. Effect of timing and intensity of grazing on the herbage quality of a Mediterranean rangeland. Journal of Animal and Feed Sciences 16, Suppl. 2: 318-322.
- Henry HAL, Cleland EE, Field CB, and Vitousek PM. 2005. Interactive effects of elevated CO2, N deposition and climate change on plant litter quality in a California annual grassland. Oecologia 142: 465-473.
- Lavergne S, Garnier E, and Debussche M. 2003. Do rock endemic and widespread plant species differ under the Leaf-Height-Seed plant ecology strategy scheme? Ecology Letters 6: 398-404.
- Lavorel S, de Bello F, Grigulis K, et al. 2011. Response of herbaceous vegetation functional diversity to decreasing land use across five sites in Europe and Israel. Israel Journal of Ecology and Evolution 57: 53-72.
- Lavorel S and Garnier E. 2002. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Functional Ecology 16: 545-556.
- Lodge GM, King KL, and Harden S. 2006. Effects of pasture treatments on detached pasture litter mass, quality, litter loss, decomposition rates, and residence time in northern New South Wales. Australian Journal of Agricultural Research 57: 1073-1085.
- Moog D, Kahmen S, and Poschlod P. 2005. Application of CSR and LHS strategies for the distinction of differently managed grasslands. Basic and Applied Ecology 6: 133-143.
- Noy-Meir I, Gutman M, and Kaplan Y. 1989. Responses of Mediterranean grassland plants to grazing and protection. Journal of Ecology 77: 290-310.
- Perevolotsky A and Seligman NG. 1998. Role of grazing in Mediterranean rangeland ecosystems. BioScience 48: 1007-1017.
- Quested H, Eriksson O, Fortunel C, and Garnier E. 2007. Plant traits relate to whole-community litter quality and decomposition following land use change. Functional Ecology 21: 1016-1026.
- Sternberg M, Gutman M, Perevolotsky A, et al. 2000. Vegetation response to grazing management in a Mediterranean herbaceous community: A functional group approach. Journal of Applied Ecology 37: 224-237.
- Sternberg M, Gutman M, Perevolotsky A, and Kigel J. 2003. Effects of grazing on soil seed bank dynamics: An approach with functional groups. Journal of Vegetation Science 14: 375-386.
- Ter Braak CJF and Šmilauer P. 2002. CANOCO for Windows 4.5. Microcomputer Power, Ithaca (NY).
- Westoby M. 1998. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil 199: 213-227.
- Westoby M. 1999. The LHS strategy scheme in relation to grazing and fire. In: Eldridge and Freudenberger (Eds). Proceedings of the VIth International Rangelands Congress; 1999 Jul 19-23; Townsville, Queensland, Australia: VI International Rangelands Congress, Inc.



 מהדורה מודפסת
מהדורה מודפסת







