
סקירות
השפעות שינוי אקלים גלובלי על מערכות אקולוגיות ימיות: תמונת מצב עולמית והשלכות על הסביבה הימית בישראל
חיים טרבס המכון הלאומי לאוקיינוגרפיה, חקר ימים ואגמים לישראל, תל-שקמונה, חיפה

כיום מרב המדענים תמימי דעים כי שינוי האקלים הגלובלי הוא עובדה שאינה ניתנת לערעור. שינוי האקלים מתבטא גם בשינויים אוקיינוגרפיים שלהם השלכות אקולוגיות מרחיקות לכת. מאמר זה סוקר את השינויים שנצפו בשנים האחרונות בחברות אקולוגיות ימיות עקב שינויי האקלים הגלובלי, ובוחן את התחזיות השונות על העתיד האקולוגי של הים. בסקירה מודגשים העלייה בטמפרטורה, בחומציות ובמפלס פני הים, ונידונות גם ההשלכות האפשריות של שינוי האקלים על האקולוגיה בימים הגובלים בישראל. מחקרים ארוכי טווח מראים הסטה של גבולות התפוצה של מינים ימיים רבים לכיוון הקטבים עקב עלייה בטמפרטורות של מי הים. במערב הים התיכון הביא אירוע של עלייה קיצונית בטמפרטורה לידי תמותה רחבת היקף של מינים רבים הקשורים לקרקעית. מבנה חברות החי על חופים סלעיים ליד אזורי פליטת מים חמים של תחנות כוח, שונה לגמרי ממבנה החברות באזורי ביקורת – עדות לשינויים הצפויים בהרכב החברות האקולוגיות עם העלייה בטמפרטורה. עלייה בחומציות הים, עקב עלייה בריכוזי הפחמן הדו-חמצני במים, גורמת לפגיעה בייצורים בוני שלד כדוגמת חלזונות וצדפות, ולפגיעה בקצב השקעת השלד של אלמוגים. מודלים חוזים התמוססות של שוניות אלמוגים שלמות אם יתממשו התחזיות השחורות בנוגע לריכוזי פחמן דו-חמצני בסוף המאה הנוכחית. עליית מי הים צפויה לפגוע בחברות האקולוגיות בעיקר באזור החוף. נתונים ראשוניים על הבדלים במגוון המינים בעומקים שונים באזור הכרית והתת-כרית הרדוד מצביעים על האפשרות שמגוון המינים ירד בצורה משמעותית על טבלאות הגידוד הסלעיות בחופי הים התיכון בישראל עקב העלייה במפלס פני הים בעשורים הקרובים. יש צורך בניטור ארוך טווח כדי להיערך לשינויים אלו.
מבוא
אין זה סוד שהאקלים בכדור הארץ משתנה, ויחד עמו האוקיינוסים. הפאנל הבין-ממשלתי לשינוי האקלים (IPCC), המונה מאות מדענים מרחבי העולם, הראה בבירור בדו"ח שפורסם בשנת 2007 כי שינויים באקלים הואצו מאמצע המאה השש-עשרה, עם המהפכה התעשייתית, והביאו לידי עלייה גלובלית בטמפרטורת האוויר והים [19]. על-פי הדו"ח, יש סיכוי של 90% שהאשמה מוטלת עלינו, בני האדם. הדו"ח מציין גם שבעשורים האחרונים התגבשו הוכחות ברורות לכך ששינויים בטמפרטורה, ב-pH (חומציות), במליחות, במפלס הים, בקרינה האולטרא-סגולה ובדגמי הזרמים באוקיינוסים, קשורים לשינוי האקלים הגלובלי המואץ. העלייה החזויה בתדירות של אירועי אקלים קיצוניים אף צפויה לגרום לעלייה בתדירות ובעוצמה של סערות קשות היכולות לפגוע בקו החוף ולהרוס שוניות אלמוגים. עד לפני שני עשורים בערך יכלו אקולוגים רק לשער מה יהיו ההשלכות של שינויים אלה על מערכות אקולוגיות בכדור הארץ [23]. כיום, לעומת זאת, ניכרות בבירור השפעות השינוי במערכות יבשתיות וימיות בכל רחבי העולם [47, 48]. סקירה שבחנה לפני שלוש שנים את ההשפעות המוכחות והאפשריות של שינוי האקלים על מערכות ימיות חופיות הראתה שהמחקר בתחום זה מתעצם במהירות [14]. עובדה זו איננה מפתיעה לנוכח העדויות הרבות על שינויים שכבר מתרחשים עקב שינוי האקלים הגלובלי. דוגמאות להשפעות אקולוגיות של העלייה בטמפרטורות באוקיינוסים הן, למשל, שינויים בפנולוגיה של אורגניזמים ימיים, הגורמים לאי-תאימות טרופית (תזונתית) בין הופעת אוכלוסיות טורפים לטרפם [7], אירועים חמורים של הלבנת אלמוגים הפוגעים בתפקוד השונית [18], הסטה לכוון הקטבים של מיני דגים שונים באוקיינוס הצפוני [26], והסטה בתפוצה של יצורים בנטוניים (החיים בקשר הדוק למצע קרקעית הים) באזורי חוף ממוזגים [15]. יש כמה דרכים לחקור את השפעת שינויי האקלים על היבטים שונים באקולוגיה: כדי לבחון השפעות אקולוגיות בקנה מידה ביו-גיאוגרפי גדול, נעשה שימוש בנתונים מסקרים כדי לבחון למשל את ההתאמה בין טמפרטורה ובין שינויים בדגמי התפוצה של מינים על קווי האורך. כדי לבחון מה יהיו ההשפעות האקולוגיות של רמות טמפרטורה ו-pH מסוימות על-פי תחזיות אקלימיות שונות, מדענים משתמשים בניסויי מעבדה מבוקרים או במערכות סגורות בשדה, בעיקר כדי לבחון השפעה על פיזיולוגיה או על שרידות של מין אחד או כמה מינים. במקרים בודדים, כפי שנראה בהמשך, אפשר להשתמש בסביבות מלאכותיות או טבעיות שכיום "מחקות" רמות צפויות של משתנים אלו כדי לבחון השפעה על מערכת אקולוגית שלמה. מודלים ביו-פיזיקליים ומודלים של מארגי מזון יכולים גם הם לשמש לבחינת השפעות על מרכיבים שונים במארג המזון.בסקירה זו נבחן כמה מן העדויות על תמורות אקולוגיות הקשורות לשינויי אקלים, ונבדוק מה אומרות תחזיות שונות על העתיד האקולוגי של הים. הדגש בסקירה יהיה על העלייה בטמפרטורה, בחומציות ובמפלס פני הים. המאמר מתבסס על פרק העוסק בנושא זה שיתפרסם בספר העומד לראות אור בקרוב,[33] אך יש בו גם ציטוטים ודוגמאות חדשות וכן דגש מיוחד על השפעות שינוי האקלים על הימים הגובלים בישראל, ובנוסף נתונים ראשוניים ומעניינים מחופינו היכולים ללמד אותנו על הצפוי למערכת זו לנוכח התחזיות הקודרות על העתיד.

השפעותיה של העלייה בטמפרטורה
הטמפרטורה היא הגורם הדומיננטי ביותר בקביעת קצבים ביולוגיים בטבע, מרמת התא ועד רמת המערכת האקולוגית כולה. לגורם זה השפעות ישירות ועקיפות על הפיזיולוגיה והאונטוגנזה (התפתחות) של אורגניזמים, על אופי האינטראקציות הטרופיות ועל עוצמתן, על המגוון הביולוגי ועל הפנולוגיה והביו-גיאוגרפיה של יצורים. לפיכך, לעלייה בטמפרטורה הקשורה לשינויי אקלים השלכות על רוב המערכות האקולוגיות הימיות, באמצעות מנגנונים שונים.ההשפעה הישירה והברורה ביותר של ההתחממות העולמית היא דרך הקשר הבסיסי ביותר בין טמפרטורה לפיזיולוגיה. תהליכים פיזיולוגיים מרכזיים, ובהם המבנה והתפקוד של חלבונים, נזילות ממברנות, תפקוד איברים [14, 16], תפקוד לב ונשימה מיטוכונדריאלית [43] מושפעים מטמפרטורת הסביבה. התאמות לתנאי הסביבה שנוצרו באורגניזמים שונים, בשילוב העובדה שרבים מהם חיים סמוך לגבול העמידות שלהם לעקת חום, עלולים להביא לידי כך שעלייה קטנה בטמפרטורה תפגע בתפקודם ואולי אפילו בשרידותם. ביטוי לכך אפשר למצוא, למשל, בתגובת חלבוני עקת חום (heat-shock proteins) בחלזונות ובסרטני כרית, החיים בתפר שבין הים ליבשה ולכן חשופים לשינויים הן בטמפרטורת האוויר והן בטמפרטורת הים [44], וכן בעלייה בהיקף המרחבי ובעוצמה של אירועי הלבנה של אלמוגים. [24] הקשר בין אירועי הלבנה לשינוי אקלים ברור [12, 18], ובמקרים רבים הוא מלווה באירועי תמותה נרחבים של אלמוגים ובשינוי כללי במבנה חברת השונית. המנגנון המכונן של אירועים אלו אינו מובן לנו לחלוטין [21], אך ידוע כי בהלבנה מאבדים האלמוגים את רוב הזואוקסנטלות – מיקרו-אצות אנדו-סימביוטיות החיות ברקמת האלמוג ואחראיות על אספקת האנרגיה לאלמוגים – וכך מקבלים האלמוגים גוון חיוור-לבן. חשיפה קצרה לטמפרטורות גבוהות תאפשר התאוששות של מינים רבים של אלמוגים, אך חשיפה של ימים רבים או שבועות, תגדיל מאוד את שיעור התמותה, ושוניות שלמות יכולות להיכחד [1]. השפעה נוספת של עלייה בטמפרטורה קשורה להשפעתה השונה על שלבים אונטוגניים (שלבי התפתחות) שונים בייצורים ימיים. עלייה זו עלולה לגרום לתזמון לקוי בין ההתפתחות הלארוולית (לארוות הן השלבים הצעירים של מינים ימיים רבים בעלי מחזור חיים מורכב, ובמקרים רבים הן פלאנקטוניות) ובין תהליכים אקולוגיים מרכזיים המבקרים את גודל האוכלוסיות, כמו אספקת מזון או עוצמת טריפה. התזמון בין אירועי רבייה לזמינות מזון לצעירים, לדוגמה, נוצר לאורך שנים רבות של אבולוציה שבמהלכן מותאמים המינים לתנאי הסביבה שלהם. פגיעה בהתאמה או בתזמון עלולה לערער את המבנה והיציבות של אוכלוסיות וחברות שונות. למשל, בצדפות החיות במשטחי הבוץ ב-Wadden Sea שבצפון אירופה נצפתה הקדמה בשחרור של תוצרי הרבייה (spawning). הלארוות הצעירות התפתחו במים לפני שיא פריחת האביב של הפיטופלנקטון הנדרש לתזונתן [29], ובשל כך סבלו מ"תת-תזונה" והתפתחותן נפגעה. כמו כן, העלייה בטמפרטורה גרמה להצטרפות מוקדמת של חסילונים צעירים, ואלה טרפו באינטנסיביות את הצדפות הצעירות והרעבות שהצטרפו זה מקרוב לאתר. לאפקט סינרגיסטי זה יכולות להיות כמובן השפעות חשובות על גודל אוכלוסיית הבוגרים. נוסף על דוגמה זו תועדו עשרות מינים אחרים שגודל אוכלוסייתם השתנה בגלל שינוי אקלימי, שינוי העלול להיות בעל השפעות חשובות על המבנה ועל היציבות של החברה האקולוגית [7].
שינויים בטמפרטורה יכולים להשפיע גם על פעילותם או על עצם קיומם של "שחקני מפתח" אקולוגיים כמו טורפים חשובים, מתחרים, מהנדסי סביבה (Ecosystem engineers) ופתוגנים. דוגמה לכך הראה סנפורד, שעקב אחר פעילות השיחור לטרף ושיעור הטריפה של כוכב הים (Pisaster ochraceus) באזור הכרית של חופיה המערביים של ארה"ב [39]. כוכב זה חי על סלעי הכרית באזור החשוף למים קרים מאוד העולים בקיץ מן העומק בשל תופעת הנביעה, תופעה הקשורה למשטר הרוחות בחוף. תצפיות וניסויים הראו כי בתקופות הקצרות שבהן שוככות הרוחות ומי השטח החמימים יותר נעים אל החוף, עולה מספר הפרטים של הכוכב וכן עולים קצבי הטריפה שלו. טענת החוקר הייתה כי עלייה משמעותית בטמפרטורה לאורך זמן בשל שינוי אקלים עלולה להביא לידי הגברת קצב הטריפה במידה שתפגע בשכיחות הטרף העיקרי שלו, צדפות גדולות היוצרות בעצמן בית גידול, ובשל כך גם במינים שונים המאכלסים את המצע שיוצרות הצדפות.
במקרים מסוימים, אירועים (טבעיים או מעשה ידי אדם) אף מזמנים לנו צפייה בניסוי "טבעי" בקנה מידה גדול, ניסוי המאפשר לנו לחזות בהשפעות אפשריות של עלייה בטמפרטורה בבתי גידול שונים. דוגמה לכך היא מעקב שביצעו כמה חוקרים אחר השפעת הפעלתה של תחנת כוח בקליפורניה על הביוטה בחוף הסלעי הסמוך [40]. החוקרים השוו את החברה האקולוגית באתר סמוך לתעלת הפליטה ממערכת הקירור לפני ואחרי הפעלת התחנה, שגרמה לעלייה בטמפרטורת המים באתר. כמו כן הושוותה חברת ביקורת המצויה בחופים מרוחקים יותר. החוקרים הראו כי מבנה החברה עבר תמורות רבות שהתבטאו בעיקר בהקטנת מספרן של כמה מיני אצות, בעיקר אצות קלפ ואצות אדומיות, שלהן חשיבות אקולוגית רבה בהיותן בעצמן בית גידול לאורגניזמים אחרים. במקביל נראתה עלייה במספרם של חסרי חוליות רועים (צמחוניים) שונים. באירוע טבעי שהתרחש בשנים האחרונות תועדה תמותה מסיבית ב-25 מינים של מאקרו-חסרי-חוליות בנטוניים (הקשורים לקרקעית) במערב הים התיכון, בעומקים הנעים בין מטרים בודדים ועד עשרות מטרים, עקב גל חום שהתרחש באירופה ב-2003 וגרם לעלייה בטמפרטורה של מי הים עד לערכים הדומים לערכים הצפויים לאזור זה בתחזיות ארוכות טווח להתחממות האוקיינוסים [11]. מובן מאליו כי אם אירועים נקודתיים אלה יהיו תכופים יותר, או אם יגיעו הטמפרטורות בקיץ לערכים אלה באופן קבוע, צפוי שינוי משמעותי במבנה החברה האקולוגית באזור זה.
תוכניות ניטור רבות שנים מראות כי עלייה בטמפרטורה גורמת לשינויים בקנה מידה ביו-גיאוגרפי נרחב. השינוי העיקרי הוא בדגמי התפוצה של מינים רבים ה"נעים" צפונה בחצי הכדור הצפוני ודרומה בחצי הכדור הדרומי, בשל עלייה בטמפרטורה באזורים הקוטביים יותר. בכך מתאפשרת הרחבת התפוצה של מינים של מים חמים על חשבונם של מינים המעדיפים מים קרים יותר. בים הצפוני, לדוגמה, נמצאה הסטה של שני שלישים מאוכלוסיות הדגים לכיוון קווי רוחב צפוניים יותר או לעומק עמודת המים, באופן שתאם את התחממות המים [28]. להסטה כזו עלולות להיות השלכות חמורות על מינים המתקיימים על גבול השרידות התרמלית שלהם. דוגמאות להסטה לכיוון הקטבים נמצאו גם באצות ובחסרי חוליות שונים. בית גידול החשוף להשפעות אלו באופן ייחודי הוא כאמור אזור הכרית (אזור שתחום בין קו המים העליון בגאות, והתחתון בעת השפל), המושפע הן מטמפרטורת המים והן מטמפרטורת האוויר, ולפיכך התפוצה הגיאוגרפית של המינים בו צפויה להצטמצם או להשתנות באופן ניכר עם העלייה בטמפרטורה. באזורים מסוימים, ניטור ארוך טווח הראה הסטה של כמה מיני כרית לכיוון הקטבים בקצב של 50 ק"מ לעשור [15].
השפעה אקולוגית חמורה של העלייה בטמפרטורה בים היא התבססות קלה יותר של מינים פולשים בבתי גידול אשר עד כה היו קרים מדי עבורם. אירועי התחממות ושינויים בדפוסי זרימת המים שמקורם באקלים הועלו במאה הקודמת בתור הגורם המרכזי המעודד פלישה של מינים טרופיים אל הים התיכון, בעיקר באגן המזרחי – אולי "הנקודה החמה" (hot spot) ביותר בעולם מבחינת פלישת מינים ימיים [34]. המסלול העיקרי לפלישה של מאות מינים לאורך חופי ישראל במאה וחמישים השנים האחרונות הוא דרך תעלת סואץ, שכרייתה חיברה את ים סוף הטרופי לים התיכון המשווני החם לאחר מיליוני שנות נתק. מאז פתיחת התעלה עברו במימיה מאות מינים, אם ישירות במים בתור לארוות, אם בתור בוגרים, אם באופן בלתי ישיר במי נטל של ספינות או כ"זקן אוניות" (fouling). קצב זה הואץ מאוד בעשורים האחרונים. תופעה זו אחראית גם להתפרצותם של כמה מינים ים-סופיים, שיוחסה בעבר לאירועי חורף חם במיוחד ששרר באזורנו, למשל בשנות החמישים, אם כי העדויות לכך היו נסיבתיות בעיקר. אפשר לשער שמינים טרופיים רבים עוברים כיום לים התיכון אך לא שורדים בטמפרטורות החורף הנמוכות מן הטמפרטורות שבים סוף, ובשל כך לא מצליחים לייסד באזור אוכלוסיות בנות-קיימא. אפשר עם זאת להעריך שמגוון המינים בחוף ישתנה מאוד אם יעלו טמפרטורות החורף המינימליות במי הים התיכון במעלה או בשתיים נוספות – עלייה אפשרית בהחלט על-פי מודלים רבים [22].
האם ייתכן שבעוד כמה עשרות שנים ייווצרו בחופינו שוניות אלמוגים? גבול הטמפרטורה התחתון של מרבית האלמוגים בוני השונית הוא 18 מעלות צלזיוס, ואילו טמפרטורות החורף בים התיכון בחוף הישראלי סובבות סביב 16–17 מעלות. עלייה במעלה אחת או בשתיים עלולה אפוא לגרום לתחילת התפתחותן של שוניות אלמוגים בחוף, אם כי עלייה גדולה בטמפרטורות המרביות בקיץ יכולה דווקא להביא לידי פגיעה באלמוגים, בשל תופעת ההלבנה, ובכך למנוע היווצרות שונית. שוניות כנראה לא יתפתחו בחופי הים התיכון למרות ההתחממות הצפויה, שכן ישנם גורמים נוספים היכולים להשפיע על התפתחות שוניות ולהגביל אותה (זמינות אור, סדימנטציה וכו'). הפן ההפוך של תחזית זו היא האפשרות של היעלמות מינים אטלנטיים החיים בחופינו, שהם גבול התפוצה המזרחי ביותר שלהם. ייתכן שחלק ממינים אלה חיים קרוב לגבול היכולת הפיזיולוגית שלהם. לפיכך, "דחיפה" קלה של טמפרטורות הקיצון כלפי מעלה יכולה להיות קטלנית עבור מינים אלה ולגרום להכחדתם במקום. מכיוון שעד היום לא נעשה ניטור מסודר של מגוון המינים ושכיחותם לאורך החוף הים-תיכוני של ישראל, ייתכן שכבר החמצנו גילויים של הכחדות כאלה.
השפעות של עלייה בחומציות (החמצת) האוקיינוסים
לאוקיינוסים תפקיד חשוב ב"אחסנת" חלק מהפחמן הדו-חמצני המצוי באטמוספרה. חישובים מראים כי האוקיינוסים אחראים גם לקליטת מחצית מכמות הפליטות מעשי ידי אדם עד שנת 1994 [37], ו-30% מהפליטות בשנים האחרונות ממש [9]. מצד שני, קליטת הפחמן הדו-חמצני צפויה לגרום לירידה של 0.5-0.3 יחידות ברמת ה-pH ב-100 השנים הקרובות במימי פני השטח של האוקיינוסים. עלייה בחומציות שכזו, אם אכן תתרחש, חורגת מכל תנודת pH מתועדת ברשומות המאובנים מ-200–300 מיליון השנים האחרונות [5]. לקצב השינוי הצפוי חשיבות גם בנוגע לסיבולת הייצורים השונים וליכולתם להסתגל לתמורות אלו. עם קצב שינוי הגדול פי מאה מהמתועד עד עתה, יכולת ההתמודדות של אורגניזמים רבים עלולה להיות מוגבלת, והשפעות ניכרות על האקולוגיה של אקוסיסטמות ימיות הן בגדר תחזית סבירה ביותר, ולמעשה כבר ניכרות בשטח.
ירידה בערכי ה-pH טומנת בחובה השלכות פיזיולוגיות על תהליכים שונים, ובהם תהליכים תת-תאיים כמו סינתזת חלבונים ושחלופי יונים [30]. או השלכות פיזיולוגיות ומכאניות ארוכות טווח, בעיקר על יצורים בוני שלד גירני (קרבונאטי), כמו ירידה בקצבים המטבוליים ובקצבי הגידול, כפי שכבר הומחש במניפולציות pH בצדפות [26]. במחקרים דומים הציגו גם קווצי-עור ורכיכות ירידה בקצבי גידול ובשרידות [41].
קצבי השקעת השלד ירדו בשנים האחרונות עקב עלייה בריכוזי פחמן דו-חמצני בחד-תאיים בעלי שלד גירני כמו קוקוליתופורידים (Coccolithophores) ופוראמניפרה (Foraminifera), וכן באצות גירניות, אלמוגים בוני שונית, וחלזונות פלאגיים מקבוצת הפטרופודה [Pteropods) [8, 9, 20, 32). ברמת הגידול של המערכת האקולוגית, תגובות אלו משפיעות בין השאר על ההרכב ועל שינויים במיני פיטופלנקטון, על-פי מידת רגישותם לשינויים בחומציות המים. נוסף על כך, שינויים כימיים אלו באוקיינוס צפויים להחליש שלדי אלמוגים קיימים ולצמצם גדילה של שוניות [18] ואף עלולים להביא לידי קריסה של אוכלוסיות פטרופודים – מרכיב מרכזי בשרשרת המזון באזור הקטבים [27]. לכך עלולות להיות השפעות על המבנה והמגוון הביולוגי במערכות אקולוגיות אלה.
גם בהיבט זה מתאפשרת לנו הצצה לעבר ההשפעות העתידיות של החמצת האוקיינוסים, בדמות נביעות געשיות טבעיות של פחמן דו-חמצני באתרים חופיים רדודים באיטליה. מחקר שהשווה את מבנה חברת הבנטוס (הקשורה לקרקעית) באזור הנביעות ובאזורים סמוכים, שבהם המים הם בעלי מידת חומציות נורמלית, הראה כי באזורים שבהם ערכי pH נמוכים בכ-0.5 יחידות מהערך הממוצע באוקיינוס כיום (ערכים החזויים עד 2100 עפ"י ה–IPCC), מספרם של יצורים משקיעי שלד גירני כמו אלמוגים, קיפודי ים ואצות גירניות, נמוכה באופן ניכר בהשוואה לאזורים עם ערכי pH נורמליים, זאת לצד עלייה ביצרנות של עשבי ים באזורים ה"חמוצים" יותר [13, 31]. כמו כן, קונכיותיהם של חלזונות סמוך לנביעות היו דקות ונקבוביות יותר מבאזורים מרוחקים.
המשמעות של ירידה בהשקעת השלד ואפילו המסה של שלדי אלמוגים עלולה להיות הרת אסון לשוניות האלמוגים בעולם. מחקר חדש מתאר ירידה של 13% בקצב השקעת השלד מאז שנת 1990 בשונית המחסום הגדולה באוסטרליה – מערכת השוניות הגדולה בעולם – והחוקרים קושרים זאת לשילוב של שני התהליכים: החמצת האוקיינוסים ועליית הטמפרטורות [6]. מודלים צופים כי עם הכפלת ריכוז הפחמן הדו-חמצני באטמוספרה, יפסיקו שוניות האלמוגים לגדול כליל, ובמקום זאת יתחילו להתמוסס [42]. מדובר בתחזית קודרת ביותר לבית הגידול הימי הנחשב העשיר ביותר במגוון המינים בסביבה הימית.
היבט מעניין נוסף הקשור לעליית ריכוז הפחמן הדו-חמצני אינו קשור בהכרח להחמצה, אלה דווקא לקצבי הגידול. הפחמן הדו-חמצני הוא מרכיב הכרחי בתהליך הגידול, והיה אפשר לשער שכמו בצמחי יבשה רבים, עלייה בריכוז הפחמן הדו-חמצני תגביר את קצבי הגידול של צמחים ימיים, וכך בעצם תפחת השפעתו באטמוספרה. אלא שבצמחים ימיים, חוץ מעשבי ים, כלומר במאקרו-אצות, הודגמה דווקא רוויה לפחמן ברוב הקבוצות, כך שאין הם צפויים להגביר את קצבי גידולן עקב העלייה בריכוזי הפחמן הדו-חמצני. לעומת זאת, ייתכן שבאזורים מסוימים, עם העלייה בריכוזי הפחמן הדו-חמצני המומס, יוחלפו מאקרו-אצות בעשבי ים, בגלל אותם הבדלים של מגבלת פחמן. הבדלים אלו נובעים מהעידנים השונים שבהם התפתחו קבוצות טקסונומיות אלו [14]. קיימים אזורים שבהם חסרים מרכיבים כימיים אחרים, החיוניים לגידול צמחים כמו ברזל במרכז האוקיינוס הדרומי. במקרה זה בודקים מדענים, כבר יותר מעשור, את האפשרות להוסיף ברזל למים כדי להגביר את ייצור הפלנקטון המיקרוסקופי הצמחי. כך אולי יהיה אפשר "לשאוב" פחמן דו-חמצני מן האטמוספרה "ולקבור" אותו בקרקעית הים – ולהפחית בכך את השפעתו כגז חממה [2, 3]. אך תוכניות אלו, בקנה המידה הגדול הנדרש לשם השפעה אמיתית על ריכוז הפחמן באטמוספירה, נשמעות כמעט דמיוניות, ויש להן מתנגדים רבים, בין השאר מכיוון שאי-אפשר לצפות אילו עוד תמורות (אולי לא רצויות) יתרחשו במערכות אקולוגיות אלו לנוכח שינויים דרמטיים כל כך בפריחת האצות.
בחופי ישראל, שהמצע הסלעי העיקרי שלהם, הכורכר, הוא מבוסס גיר, עלולה החמצת האוקיינוסים להשפיע על המצע עצמו. הדבר יכול לגרום למשל לבליה מואצת של סלעי החוף ולקריסת טבלאות הגידוד הייחודיות לחופי הארץ, שהן טבלאות שטוחות של כורכר בגובה פני הים באזור הכרית. להחמצה יכולה כמובן להיות גם השפעה על הייצורים הגירניים בבית הגידול, ובמיוחד למהנדסי הסביבה, המעצבים ומאפשרים למעשה את קיומן של הטבלאות בעוצרם את בליית הסלע בדיוק בגובה מפלס פני הים: אלה הם החלזונות השילשולניים קבועי המקום, היוצרים צברים צפופים ושומרים בגופם הגירני הקשה על הטבלאות מפני בליה [38, 45]. אך מתברר שעוד לפני שהחומציות השיגה אותם, חלזונות אלו כבר נעלמים מחופינו בשנים האחרונות, מסיבות שעדיין אינן ידועות (ואולי קשורות לשינוי האקלים). עדויות ראשוניות לכך נצפו כבר בשנות התשעים [33], וכיום הן מתחזקות: בתצפיות בשנת 2009 עדיין לא מצאנו אף לא פרט אחד של המין Dendropoma petraeum, המתקיים בעיקר בשולי טבלת הגידוד ויוצר כרכוב מוגבה. ייתכן כי המין נמצא על סף היכחדות לאורך החוף הישראלי (רילוב, נתונים שלא פורסמו). משמעות היעלמותם של החלזונות יכולה להיות לפיכך בלייה מהירה במיוחד של הסלע וקריסת הטבלאות בתוך כמה עשרות או מאות שנים, תלוי בקצב הבלייה הטבעי ובהשפעתו של תהליך החמצת האוקיינוסים.
עליית מפלס פני הים
אחת ההשפעות של העלייה בטמפרטורה באטמוספרה ובים היא עלייה במפלס פני הים. הסיבות לכך מגוונות, אך העיקריות שבהן הן המסת הקרחונים המכסים את היבשות והתפשטות תרמלית של נפח המים [4, 25]. שינוי במשטר הזרמים באוקיינוסים יכול גם הוא לתרום לעליית המפלס במקומות מסוימים. המודלים האוקייניים מראים כי מפלס הים יעלה במידה שונה במקומות שונים בעולם. רוב התחזיות מדברות על עלייה של עשרות סנטימטרים במאה הנוכחית, ויש כאלה החוזים אף עלייה במטרים ספורים. המשמעות של עליית מפלס פני הים לאוכלוסייה האנושית ברורה ביותר: היא תגרום להצפת אזורים גדולים של החוף, להמלחת מי התהום ולהאצה של הבלייה בחוף, במיוחד לנוכח הצפי להתגברות העוצמה והתדירות של סערות בים. כמה איים באוקיינוס השקט שמרבית שטחם מצוי רק מעט מעל הגובה הממוצע של פני הים, כבר חווים זאת הלכה למעשה, ויש סיכוי רב שתושביהם יצטרכו להיערך לקראת פינוי בשנים הקרובות.

ההשפעות האקולוגית של עליית מפלס הים מורכבות יותר, וגם הן תלויות מאוד באזור. מטבע הדברים, המערכות האקולוגיות שיושפעו ביותר משינויים אלו הן של הכרית והתת-כרית הרדוד. להערכת Galbraith, גם על-פי תחזיות שמרניות של עליית מפלס הים, יאבדו 70–20 אחוזים משטחי הכרית בכמה מהמפרצים הגדולים ביותר בארה"ב, שבהם אוכלוסיות גדולות של עופות חופים ועופות ים חורפים [10]. אובדן כזה יכול להקטין באופן משמעותי את גודל אוכלוסיותיהן של עופות אלו, ובכך עלול להשפיע על כלל מארג המזון של בית הגידול. בחופים שבהם משרעת הכרית היא מינימלית, השפעתה של עליית המפלס, ואפילו קטנה, צפויה להיות משמעותית ביותר. דוגמה לכך היא החוף הישראלי של הים התיכון. בחופינו משרעת הגאות והשפל היא 30–20 ס"מ בממוצע, והמקסימום הוא כ-40 ס"מ. על-פי נתונים שאוסף כבר שנים רבות דב רוזן מהמכון הלאומי לאוקיינוגרפיה של חקר ימים ואגמים לישראל, עלה המפלס משנת 1992 ועד 2008 ב-8.5 ס"מ [36], כלומר כשליש מן המשרעת הממוצעת – ערך יחסי גבוה מאוד לכל הדעות. רוזן מציין שיש אף תחזיות לעלייה של כמטר שלם בחופינו, עוד במאה הזאת. מעבר לבעיות שעלייה כזו יכולה ליצור עבור האדם לאורך החוף (הצפות, בליה וקריסה של צוקים ועוד), המשמעות לבית הגידול הייחודי של הכרית בישראל, טבלאות הגידוד, עלולה להיות הרת אסון. למרות משרעת הגאות והשפל המינימלית בחופי ישראל, אזור הכרית בחופינו נרחב ביותר. הסיבה לכך היא שהטבלאות המצויות בדיוק בגובה מפלס פני הים, וחשופות לסירוגין לאוויר (בזמן השפל) ולמי הים (בזמן הגאות או פעילות גלים), יכולות להגיע במקומות מסוימים לרוחב של עשרות מטרים, ומתקיים עליהן מגוון מינים ייחודי שאינו קיים על משטחים סלעיים במפלסים שמעליהן או מתחתיהן. סקר עדכני בחוף שקמונה מראה כי עושר המינים על פני הטבלה גבוה פי שניים-שלושה ממפלסים נמוכים ממנו ב-10–50 ס"מ בלבד, כלומר בחגורות גובה הגובלות באזור התת-כרית או מצויות בו ממש (איור 1). משמעות הדבר ברורה: עלייה של עשרה ס"מ ויותר עלולה לגרום לצניחה של מגוון המינים לאורך החוף, מכיוון שלטבלאות נרחבות אלו אין תחליף דומה במפלסים גבוהים יותר. גב הטבלאות ברוב אזורי החוף מאופיין בחוף חולי (כמו בשקמונה לדוגמה, איור 2) או בקירות אנכיים בגובה מטרים ספורים (לדוגמה חוף הבונים ואכזיב). כלומר, לא קיימים משטחים סלעיים גבוהים שיכולים להוות חלופה לטבלאות הגידוד, ואלה גם לא ייווצרו בעתיד אם החלזונות בוני טבלאות הגידוד אכן נכחדים באזורנו. נוסף על כך, ידוע שחברות החי והצומח באזור הכרית שונים מאוד בין מפנים אנכיים ואופקיים, ולפיכך, גם בשל סיבה זו, אופי מגוון המינים יהיה שונה מאוד אם יעלה מפלס הים בחופינו [46].
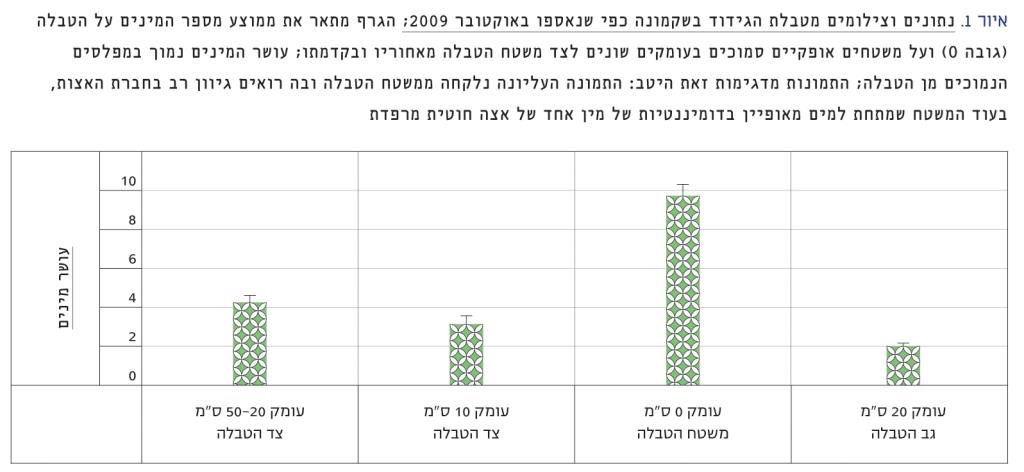
איור 1
נתונים וצילומים מטבלת הגידוד בשקמונה כפי שנאספו באוקטובר 2009
הגרף מתאר את ממוצע מספר המינים על הטבלה (גובה 0) ועל משטחים אופקיים סמוכים בעומקים שונים לצד משטח הטבלה מאחוריו ובקדמתו; עושר המינים נמוך במפלסים הנמוכים מן הטבלה; התמונות מדגימות זאת היטב: התמונה העליונה נלקחה ממשטח הטבלה ובה רואים גיוון רב בחברת האצות, בעוד המשטח שמתחת למים מאופיין בדומיננטיות של מין אחד של אצה חוטית מרפדת

איור 2
חוף חולי/חלוקי בגב טבלאות הגידוד בשקמונה (צילום עליון); קירות אנכיים בגב טבלאות הגידוד בשמורת דור-הבונים (צילום תחתון)
סיכום
אפשר לומר בוודאות כי שינוי האקלים הגלובלי כבר הטביע את חותמו על האקולוגיה של הים ברחבי העולם, והוא משנה בקצב גובר והולך מגוון רחב של מערכות אקולוגיות. התחזיות לעתיד קודרות למדי: השילוב של העלייה בטמפרטורה, במפלס פני הים ובחמיצות המים, נוסף על שינוי הזרמים, תדירותן ועוצמתן של סערות ועוד, ישנה את האקולוגיה באוקיינוסים באופן דרמטי. אך הבנתנו בנוגע למלוא משמעותם של שינויים אלו עדיין בחיתוליה, ונדרש עוד מחקר רב בתחום. גם החוף הישראלי, הן שונית האלמוגים באילת והן הים תיכון, עוברים ועוד צפויים לעבור שינויים דרמטיים בעשרות השנים הקרובות. לנוכח זאת, מדענים ואנשי ניהול ומדיניות חייבים לקיים דיאלוג שמטרתו לגבש תוכניות ניטור ארוכות טווח, וכך גם מחקר בסיסי על התהליכים בבתי גידול אלו ועל האופן שבו יגיבו לשינויים הצפויים. מהלכים אלו יאפשרו לעקוב אחר התמורות ולבנות תחזיות ריאליות באשר לכיוונים שבהם צפויה להשתנות המערכת הימית בטווח הזמן הנראה לעין. במקביל, יש להיערך לשינויים אלו על-ידי תכנון כלים לניהול יעיל על בסיס מדעי, אך אין ספק כי הפעילות החשובה ביותר יכולה להיות בתרומתנו למניעת המשך ההידרדרות העולמית, באמצעות השתתפות במאמצים לבלימת הגורמים היוצרים את ההתחממות, בעיקר פליטת גזי חממה.
בימים אלו מנסה קבוצת המחקר שלנו במכון לחקר ימים ואגמים בחיפה לבנות בסיס לתוכנית ניטור כזו באזור החוף הסלעי של הים התיכון, תוכנית שתאפשר מעקב צמוד אחר שפיעות המינים בבית גידול זה. תוכנית הניטור מיועדת לתעד שינויים במבנה חברת החי והצומח – קריסה והיעלמות של מינים מקומיים וכן הופעת מינים חדשים לאורך החוף, ולקשור שינויים אלו למגמות בתנאי הסביבה. כמו כן יתבצעו מחקרים בים ובמעבדה שיסייעו בחיזוי חלק מהשינויים הצפויים למינים רגישים ולחברה האקולוגית כולה.
מקורות
- Baker AC, Glynn PW, and Riegl B. 2008. Climate change and coral reef bleaching: An ecological assessment of long-term impacts, recovery trends and future outlook. Estuarine Coastal and Shelf Science 80:435-471.
- Behrenfeld MJ, Bale AJ, Kolber ZS, Aiken J, and Falkowski PG. 1996. Confirmation of iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean. Nature 383: 508-511.
- Boyd PW, Watson AJ, Law CS, Abraham ER, Trull T, Murdoch R, Bakker DCE, Bowie AR, Buesseler KO, Chang H, Charette M, Croot P, Downing K, Frew R, Gall M, Hadfield M, Hall J, Harvey M, Jameson G, LaRoche J, Liddicoat M, Ling R, Maldonado MT, McKay RM, Nodder S, Pickmere S, Pridmore R, Rintoul S, Safi K, Sutton P, Strzepek R, Tanneberger K, Turner S, Waite A, and Zeldis J. 2000. A mesoscale phytoplankton bloom in the polar Southern Ocean stimulated by iron fertilization. Nature 407: 695-702.
- Cabanes C, Cazenave A, and Le Provost C. 2001. Sea level rise during past 40 years determined from satellite and in situ observations. Science 294: 840-842.
- Caldeira K and Wickett ME. 2003. Anthropogenic carbon and ocean pH. Nature 425: 365-365.
- De'ath G, Lough JM and Fabricius KE. 2009. Declining Coral Calcification on the Great Barrier Reef. Science 323: 116-119.
- Edwards M and Richardson AJ. 2004. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 430: 881-884.
- Fabry VJ. 2008. Ocean science – Marine calcifiers in a high-CO2 ocean. Science 320: 1020-1022.
- Feely RA, Sabine CL, Lee K, Berelson W, Kleypas J, Fabry VJ and Millero FJ. 2004. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 305: 362-366.
- Galbraith H, Jones R, Park R, Clough J, Herrod-Julius S, Harrington B, and Page G. 2002. Global climate change and sea level rise: Potential losses of intertidal habitat for shorebirds. Waterbirds 25: 173-183.
- Garrabou J, Coma R, Bensoussan N, Bally M, Chevaldonne P, Cigliano M, Diaz D, Harmelin JG, Gambi MC, Kersting DK, Ledoux JB, Lejeusne C, Linares C, Marschal C, Perez T, Ribes M, Romano JC, Serrano E, Teixido N, Torrents O, Zabala M, Zuberer F, and Cerrano C. 2009. Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Global Change Biology 15: 1090-1103.
- Glynn PW. 1993. Coral reef bleaching: ecological perspectives. Coral Reefs 12: 1-17.
- Hall-Spencer JM, Rodolfo-Metalpa R, Martin S, Ransome E, Fine M, Turner SM, Rowley SJ, Tedesco D, and Buia MC. 2008. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 454: 96-99.
- Harley CDG, Hughes AR, Hultgren KM, Miner BG, Sorte CJB, Thornber CS, Rodriguez LF, Tomanek L, and Williams SL. 2006. The impacts of climate change in coastal marine systems. Ecology Letter 9: 228-241.
- Helmuth B, Mieszkowska N, Moore P, and Hawkins SJ. 2006. Living on the edge of two changing worlds: Forecasting the responses of rocky intertidal ecosystems to climate change. Annual Review of Ecology Evolution and Systematics 37: 373-404.
- Hochachka PW and Somero GN. 2002. Biochemical adaptation: mechanism and process in physiological evolution. Biochemical adaptation: mechanism and process in physiological evolution. i-xi, 1-466.
- Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R, Muthiga N, Bradbury RH, Dubi A, and Hatziolos ME. 2007. Coral reefs under rapid climate change and ocean acidification. Science 318: 1737-1742.
- Hughes TP, Baird AH, Bellwood DR, Card M, Connolly SR, Folke C, Grosberg R, Hoegh-Guldberg O, Jackson JBC, Kleypas J, Lough JM, Marshall P, Nystrom M, Palumbi SR, Pandolfi JM, Rosen B, and Roughgarden J. 2003. Climate change, human impacts, and the resilience of coral reefs. Science 301: 929-933.
- IPCC 2007. Intergovernmental Panel on Climate Change Report – Technical Summary. In p. 74.
- Kleypas JA, Buddemeier RW, Archer D, Gattuso JP, Langdon C, and Opdyke BN. 1999. Geochemical consequences of increased atmospheric carbon dioxide on coral reefs. Science 284: 118-120.
- Lesser MP. 2006. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annual Review of Physiology 68: 253-278.
- Lipkin Y and Safriel U. 1971. Intertidal zonation of the rocky shores at Mikhmoret (Mediterranean, Israel). Journal of Ecology 59: 1-30.
- Lubchenco J, Navarrete SA, Tissot BN, and Castilla JC. 1993. Possible ecological responses to global climate change: nearshore benthic biota of northeastern Pacific coastal ecosystems. In: Earth system responses to global change (Eds. Mooney HA, Fuentes ER, and Kronberg BI). Academic Press, Inc., San Diego, CA.
- McWilliams JP, Cote IM, Gill JA, Sutherland WJ, and Watkinson AR. 2005. Accelerating impacts of temperature-induced coral bleaching in the Caribbean. Ecology 86: 2055-2060.
- Meehl GA, Washington WM, Collins WD, Arblaster JM, Hu AX, Buja LE, Strand WG, and Teng HY. 2005. How much more global warming and sea level rise? Science 307: 1769-1772.
- Michaelidis B, Ouzounis C, Paleras A, and Portner HO. 2005. Effects of long-term moderate hypercapnia on acid-base balance and growth rate in marine mussels Mytilus galloprovincialis. Marine Ecology-Progress Series 293: 109-118.
- Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, Feely RA, Gnanadesikan A, Gruber N, Ishida A, Joos F, Key RM, Lindsay K, Maier-Reimer E, Matear R, Monfray P, Mouchet A, Najjar RG, Plattner GK, Rodgers KB, Sabine CL, Sarmiento JL, Schlitzer R, Slater RD, Totterdell IJ, Weirig MF, Yamanaka Y, and Yool A. 2005. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437: 681-686.
- Perry AL, Low PJ, Ellis JR, and Reynolds JD. 2005. Climate change and distribution shifts in marine fishes. Science 308: 1912-1915.
- Philippart CJM, van Aken HM, Beukema JJ, Bos OG, Cadee GC, and Dekker R. 2003. Climate-related changes in recruitment of the bivalve Macoma balthica. Limnology and Oceanography 48: 2171-2185.
- Portner HO, Langenbuch M, and Michaelidis B. 2005. Synergistic effects of temperature extremes, hypoxia, and increases in CO2 on marine animals: From Earth history to global change. Journal of Geophysical Research-Oceans 110.
- Riebesell U. 2008) Climate change – Acid test for marine biodiversity. Nature 454: 46-47.
- Riebesell U., Zondervan I., Rost B., Tortell P.D., Zeebe R.E. and Morel FMM. (2000) Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 407, 364-367.
- Rilov G, Benayahu Y, and Gasith A. 2004. Prolonged lag in population outbreak of an invasive mussel: a shifting-habitat model. Biological Invasions 6: 347-364.
- Rilov G and Galil B. 2009. Marine bioinvasions in the Mediterranean Sea – history, distribution and ecology. In: Biological Invasions in Marine Ecosystems: Ecological, Management, and Geographic Perspectives (Eds. Rilov G and Crooks JA), pp. 3-11. Springer-Verlag, Heidelberg, Germany.
- Rilov G, and Treves H. 2009. Climate change effects on marine ecological communities. In: Seaweeds and their role in globally changing environments (Eds. Israel A, Rachel E, and Joseph S). Springer.
- Rosen D. 2008. Monitoring boundary conditions at Mediterranean Basin − Key element for reliable assessment of climate change, variability and impacts at Mediterranean basin shores. In: Towards an integrated system of Mediterranean marine observatories, pp. 107-111. CISEM workshop, La Spezia.
- Sabine CL, Feely RA, Gruber N, Key RM, Lee K, Bullister JL, Wanninkhof R, Wong CS, Wallace DWR, Tilbrook B, Millero FJ, Peng TH, Kozyr A, Ono T, and Rios AF. 2004. The oceanic sink for anthropogenic CO2. Science 305: 367-371.
- Safriel UN. 1974. Vermetid gastropods and intertidal reefs in Israel and Bermuda. Science 186: 1113-1115.
- Sanford E. 1999. Regulation of keystone predation by small changes in ocean temperature. Science 283: 2095-2097.
- Schiel DR, Steinbeck JR, and Foster M. 2004. Ten years of induced ocean warming causes comprehensive changes in marine benthic communities. Ecology 85: 1833-1839.
- Shirayama Y and Thornton H. 2005. Effect of increased atmospheric CO2 on shallow water marine benthos. Journal of Geophysical Research-Oceans 110.
- Silverman J, Lazar B, Cao L, Caldeira Kand Erez J. 2009. Coral reefs may start dissolving when atmospheric CO2 doubles. Geophysical Research Letters 36.
- Somero GN. 2002. Thermal limits to life: underlying mechanisms and adaptive plasticity. Integrative and Comparative Biology 42: 1316-1316.
- TomanekL and Somero GN. 1999. Evolutionary and acclimation-induced variation in the heat-shock responses of congeneric marine snails (genus Tegula) from different thermal habitats: Implications for limits of thermotolerance and biogeography. Journal of Experimental Biology 202: 2925-2936.
- Tzur Y and Safriel UN. 1978. Vermetid platforms as indicators of coastal movement. Israel Journal of Earth Sciences 124-127.
- Vaselli S, Bertocci, Maggi E, and Benedetti-Cecchi L. 2008. Assessing the consequences of sea level rise: effects of changes in the slope of the substratum on sessile assemblages of rocky seashores. Marine Ecology-Progress Series 368: 9-22.
- Walther GR, Berger S, and Sykes MT. 2005. An ecological 'footprint' of climate change. Proceedings of the Royal Society B-Biological Sciences 272: 1427-1432.
- Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, and Bairlein F. 2002. Ecological responses to recent climate change. Nature 416: 389-395.



 מהדורה מודפסת
מהדורה מודפסת








