
חזית המחקר
השפעות קצרות טווח של עוצמות רעייה שונות על הביומסה ועל מגוון המינים העשבוני ביער איקליפטוס נטוע בצפון-מערב הנגב
דני ברקאי מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקני
יעקב קנול מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקני
ירון זיו המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגב ( [email protected])
גיא רותם המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגב ([email protected])
אלי צעדי המחלקה למשאבי טבע, המכון למדעי הצמח ומרכז מחקר גילת, מִנהל המחקר החקלאי – מרכז וולקני ([email protected])

עדר צאן רועה ביער נחל אסף, פברואר 2015 | צילום: אמיר גליק
אמיר גליק
מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקני; המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגבדני ברקאי
מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקנייעקב קנול
מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקניירון זיו
המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגבגיא רותם
המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגבאלי צעדי
המחלקה למשאבי טבע, המכון למדעי הצמח ומרכז מחקר גילת, מִנהל המחקר החקלאי – מרכז וולקניאמיר גליק
מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקני; המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגבדני ברקאי
מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקנייעקב קנול
מרכז מחקר גילת, מִִנהל המחקר החקלאי – מרכז וולקניירון זיו
המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגבגיא רותם
המעבדה לאקולוגיה מרחבית, המחלקה למדעי החיים, אוניברסיטת בן-גוריון בנגבאלי צעדי
המחלקה למשאבי טבע, המכון למדעי הצמח ומרכז מחקר גילת, מִנהל המחקר החקלאי – מרכז וולקנילרעיית מקנה מיוחסים יתרונות רבים, ובהם הפחתת ביומסה עשבונית הגורמת להיווצרות שרפות והעלאת מגוון המינים. עקב כך, קק"ל יוזמת רעיית עדרי צאן ביערותיה כדי למנוע הצטברות חומר דליק. מחקר ראשוני זה בחן כיצד עוצמות רעייה שונות משפיעות בטווח הקצר על הביומסה ועל מגוון המינים העשבוני ביער נחל אסף בצפון-מערב הנגב. במסגרת זו בוצע מחקר קצר טווח שבמהלכו נחשפו אזורים שונים ביער לטיפולי רעייה בעוצמות שונות ובמרחקים שונים ממכלאת הכבשים. כמו כן, בוצע מעקב אחר הביומסה ומגוון המינים בעונת הרעייה. המחקר מצא כי הביומסה העשבונית עלתה בחלקות ללא רעייה, אך לא השתנתה באופן מובהק תחת טיפולי הרעייה. נוסף על כך, נמצא כי בעוצמת רעייה בינונית עלה מגוון המינים העשבוניים באופן הבולט ביותר בהשוואה לשאר עוצמות הרעייה. התוצאות הללו מראות כי עוצמות רעייה בינוניות, כלומר כאשר הצמחייה העשבונית חשופה לרעייה לסירוגין ולא לזמן ממושך, הן אכן עוצמות הרעייה שיכולות לענות על הצורך בהפחתת הביומסה העשבונית למניעת שרפות ולשמירה על מגוון המינים ביערות. נוסף על כך, לא מומלץ למנוע רעייה ביערות הנטועים, שכן מחקר זה הראה כי מניעת רעייה מגדילה את הביומסה הדליקה, ומונעת עלייה במגוון המינים.
על קצה המזלג
- לממשק רעיית צאן מיוחסות השפעות חיוביות על המערכת הצמחית, כגון הפחתת סכנת השרפות והגדלת המגוון הביולוגי.
- על אף הידע הרב בנושא השפעות רעייה על מערכות צומח אחרות )עשבוניות וחורש ים תיכוני( יש פער ידע באשר להשפעות רעייה בשטחים שניטעו בהם עצי איקליפטוס.
- שכיחותם של יערות איקליפטוס נטועים בצפון- מערב הנגב מצריכה גיבוש גישה שתאפשר קיום מגוון מינים גבוה, אך גם תמנע התפרצות שרפות והתפשטותן.
- המאמר מספק בסיס ראשוני לקביעת מדיניות לשילוב רעיית עדרים במערכות צומח ביער איקליפטוס נטוע, כולל קביעת משטר הרעייה המיטבי ביער, שישלב מניעת שרפות עם קיום מרבי של מגוון מיני צומח.
המערכת
הלכה למעשה
מבוא
רעיית מקנה היא שימוש קרקע נפוץ ברחבי העולם, ויש לה השפעות מרחיקות לכת על מרכיבים ביוטיים ואביוטיים של מערכות אקולוגיות [2, 4, 24]. אלפי שנים של רעיית מקנה והפרעות אנושיות גרמו לחברת הצומח באזור מזרח הים התיכון לפתח תכונות שמקנות לה עמידות לרעייה [11, 19]. יתרה מכך, מדענים מאמינים כי חברות צומח באזורים אלה עברו ברִירה (סלקציה) חזקה, ומיני הצמחים בהן תלויים בהפרעות רעייה [17, 19, 23]. ואכן, מחקרים רבים תומכים ברעיון שניהול נכון של רעיית מקנה ובקר יכול להעלות את מגוון מיני הצומח [13, 16, 18].
מלבד השימושים המסורתיים במקנה למזון ולמחיה, גופים המנהלים שטחים פתוחים נעזרים ברעיית מקנה ככלי ניהול להפחתת עשבייה דליקה בתת-היער [7]. בישראל נפוצות מאוד שרפות יער הנגרמות ממקור אנושי [10], ולכן קק"ל משלבת רעיית מקנה ביערותיה כדי להפחית את תדירות השרפות ואת חומרתן [1, 5].
השערת ההפרעה הבינונית [3] גורסת כי מגוון המינים המרבי צפוי להימצא תחת רמות הפרעה בינוניות (או עוצמת רעייה בינונית), שההפרעה בהן לא חזקה מספיק כדי לגרום להכחדה, ולא חלשה מדי כך שרמות התחרות מאפשרות קיום של מינים רבים. עם זאת, למגוון המינים קשר הדוק עם יצרנות ראשונית [8, 20]. מגוון מינים צפוי להראות תגובה אונימודלית ליצרנות [21], כלומר התפלגות בעלת נקודת מקסימום אחת, עם מגוון מינים מרבי ברמות יצרנות בינוניות [8, 20]. בהתבסס על מערכת היחסים בין יצרנות ראשונית למגוון מינים, מחקרים רבים תומכים ברעיון כי השפעת רעייה על מגוון המינים משתנה לאורך שיפוע היצרנות [9, 15, 20]. במאמר סקירה שבחן השפעות רעייה על מגוון מינים באזורים ברמות יצרנות שונות [20], הוצעה השערת היפוך הרעייה (Grazing Reversal), שלפיה באזורים עם רמת יצרנות נמוכה, רעייה מקטינה את מגוון המינים עקב הגבלת משאבים שמונעת צמיחה מחדש. לעומת זאת, באזורים עם יצרנות גבוהה, רעייה מגבירה דומיננטיות של מינים נדירים ובלתי אכילים, מה שמניב מגוון גדול יותר. ההשפעה ההדדית הזו בין רעייה, מגוון מינים ויצרנות מתרחשת גם בקני מידה קטנים לאורך אזורי מרעה שונים [15].
בעקבות השערת היפוך הרעייה בוחן מודל הרעייה הכללי [9] את השפעות הרעייה על מגוון צומח באתרים עם רמת יצרנות שונה (צחיח למחצה לעומת ים תיכוני) ועם היסטוריות רעייה שונות (ארוכה לעומת קצרה). המודל מציע כי באזורים עם רמת יצרנות נמוכה והיסטוריית רעייה ארוכה (כמו מזרח הים התיכון), מגוון המינים יורד במתינות ככל שעוצמת הרעייה מתגברת, כפי שמוצע בהשערת היפוך הרעייה. לעומת זאת, אזורים המתאפיינים ביצרנות גבוהה (כלומר בעלי משטר גשמים משמעותי יותר) ובהיסטוריית רעייה ארוכה צפויים להראות תגובה אונימודלית של מגוון מינים לעוצמת רעייה, בדומה להשערת ההפרעה הבינונית [3].
לסיכום, השערת היפוך הרעייה ומודל הרעייה הכללי (עבור אזורים עם יצרנות נמוכה והיסטוריית רעייה ארוכה) חוזים תגובה לינארית שלילית של מגוון מינים כפונקציה של עוצמת הרעייה. מנגד, השערת ההפרעה הבינונית ומודל הרעייה הכללי (עבור אזורים ים תיכוניים והיסטוריית רעייה ארוכה) צופים תגובה אונימודלית. התחזיות הסותרות הללו מראות כי אין הסבר ברור לדרך ההשפעה של הרעייה על מגוון מיני הצומח במערכות אקולוגיות שונות, ולכן נחוצים מחקרים נוספים שיתרמו להבנת המנגנונים הללו.

רעייה משפיעה לא רק על מגוון המינים, אלא גם על הרכב חברת הצומח. מחקרים רבים נערכו כדי לבדוק כיצד קבוצות תפקודיות של צמחים, כלומר קבוצות מיני צמחים בעלות מאפיין משותף, כגון צורת חיים או השתייכות לאותה משפחה, מגיבות לעוצמות רעייה שונות [4, 7, 12, 18]. באופן כללי, קבוצות צמחים שונות מראות עלייה, ירידה או אי-שינוי בשפע בתגובה לעוצמות רעייה שונות [12]. באזורים יובשניים, צמחים חד-שנתיים, שרועים ונמוכים נפוצים יותר באזורים שיש בהם רעייה, בשל עמידותם לרעייה ויכולותיהם להימנע מרעייה [4, 22]. מצד שני, צמחים רב-שנתיים, גבוהים וטעימים (palatable) מוגדרים כרגישים לרעייה, ולכן מראים שפע נמוך יותר באזורים שיש בהם רעייה. בישראל בוצעו מחקרים הבודקים את השפעות הרעייה על קבוצות צמחים שונות בכרי המרעה בצפון הארץ [12, 22] ובחורש הים התיכוני [7], אך לא ביערות נטועים באזורים הצחיחים למחצה.
מחקר זה נועד לבדוק את ההשפעות קצרות הטווח של רעיית צאן על ביומסה ועל מגוון מינים עשבוניים ביער איקליפטוס נטוע בצפון-מערב הנגב. מטרת המחקר היא למצוא את הקשר שבין ביומסה למגוון מינים עשבוניים בתת-היער, בתקווה למצוא משטר רעייה מתאים, שמפיק רמות ביומסה נמוכות ומגוון גבוה. ישנם מעט מחקרים הבוחנים את ההשפעות קצרות הטווח של רעייה על צמחיה ביערות איקליפטוס נטועים בצפון הנגב, ולכן למחקר זה חשיבות גבוהה לממשק יערות אלה.
שיטות
שטח המחקר
אתר המחקר מוקם ביער נחל אסף (31°20’N, 34°25’E; איור 1), יער איקליפטוס נטוע בצפון-מערב הנגב. האתר ממוקם באזור מעבר בין אקלים ים תיכוני למדברי. כמות הגשם הממוצעת לשנים 1980–2010 הייתה 311 מ"מ (השירות המטאורולוגי). בשנת המחקר 2014–2015 כמות הגשם הייתה 437 מ"מ, משמע שנה גשומה. ביער ניטעו ב-1986 עצי איקליפטוס המקור (Eucalyptus camaldulensis) והם העצים היחידים שגדלים בו. במשך החורף מכסה צמחייה עשבונית עשירה (חד-שנתיים בעיקר) את תת-היער. העשבוניים מגיעים לשיא הביומסה לקראת האביב, באפריל.

איור 1
אזור המחקר
מיקום שטח המחקר (באדום מימין) ותצלום לוויין עם מיקום הגדורות, המכלאה ביער (שמאל) ומכלאת הצאן.
עיצוב הניסוי
עדר הניסוי מורכב מכ-90 כבשים מזן אוואסי ומכ-30 עזים בלָדיות. רעיית העדר החלה בפברואר 2015 ונמשכה עד אפריל 2015. שמונה מערכים של גדורות בשטח 15X5 מטרים כל אחת נבנו ביער במרחקים שונים ממכלאת הכבשים (100, 200, 300, 400, 500, 600, 800 ו-1,000 מטרים). כדי לשלוט בעוצמות הרעייה חולקה כל גדורה לשלושה טיפולים (5X5 מטר): הגדורה האמצעית הייתה סגורה במשך כל הניסוי (ללא רעייה); גדורה פינתית אחת הייתה פתוחה לסירוגין במשך שבוע ואז סגורה שבועיים, כך במשך חודשיים (עוצמת רעייה נמוכה); גדורה פינתית שנייה הייתה פתוחה במשך שבועיים ואז סגורה שבוע, וזאת לסירוגין במשך חודשיים (עוצמת רעייה בינונית); לצד שלוש הגדורות סומן שטח לא מגודר של 5X5 מטרים שהעדר יכול לרעות בו ללא הפרעה (עוצמת רעייה גבוהה).
דגימת צמחייה
הצמחייה נדגמה פעמיים בכל אחת מ-32 החלקות: בתחילת פברואר 2015 (לפני תחילת הרעייה) ובאמצע אפריל 2015 (לאחר הרעייה). בתוך כל גדורה נאספה צמחייה מאזורים מסומנים מחמש מסגרות ריבועיות בגודל 20X20 ס"מ. לאחר עקירת העשבים מוינו הצמחים וזוהו במעבדה, ולאחר ייבוש בתנור הם נשקלו. מגוון מינים חושב לפי מדד פישר למגוון מינים [6]. הצמחים מוינו לחמש קבוצות תפקודיות לפי גובה, מחזור חיים, טעימוּת (palatability) וטקסונומיה, בהתאם ל- Noy-Meir[12] ול-Sternberg [22] : דגניים נמוכים (משפחת הדגניים Poaceaea, מתחת ל-50 ס"מ בבגרות), דגניים גבוהים (משפחת הדגניים Poaceae, מעל 50 ס"מ בבגרות), קטניות (משפחת הפרפרניים Fabaceae), קוצניים (משפחת המורכבים Asteraceaeוקוצניים ממשפחות אחרות) ורחבי עלים (שאר הדו-פסיגיים).
ניתוח סטטיסטי
הנתונים משתי הדגימות נותחו באמצעות repeated measures ANOVA כדי לבחון את השפעת עוצמת הרעייה על הביומסה ועל מגוון מיני העשבוניים. המבחנים נעשו בתוכנת STATISTICA 10 (StatSoft, Tulsa, OK, USA, 2011). נוסף על כך, כדי לבדוק את השתנות הרכב המינים, יצרנו טבלת מרחקים באמצעות מדד Bray-Curtis, ואת תוצאותיו שיבצנו במרחב דו-ממדי בניתוח nMDS עם תוכנת Primer 6.
תוצאות
בסך הכול זוהו כ-80 מיני עשבוניים ביער. דגניים (גבוהים ונמוכים יחד) היו הקבוצה הדומיננטית עם 63% מכלל העשבים. הקבוצה הדומיננטית השנייה הייתה רחבי העלים עם 15%, ואחריהם הקוצניים עם 12%. טבלה 1 מפרטת את 20 המינים הנפוצים ביותר ביער לאורך הניסוי.
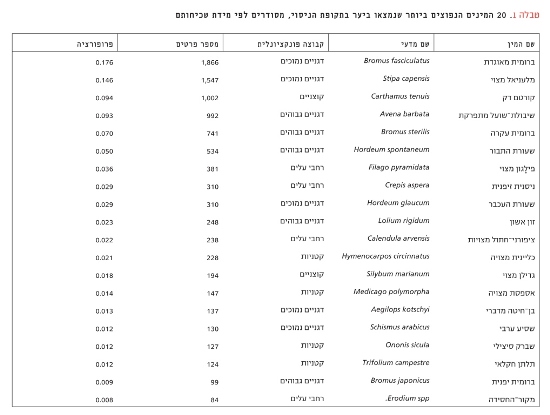
טבלה 1
20 המינים הנפוצים ביותר שנמצאו ביער בתקופת הניסוי, מסודרים לפי מידת שכיחותם
ביומסה
ביער הוותיק הראה מבחן ANOVA עבור ביומסה לאורך שני אירועי דגימה וטיפולי רעייה שונים הבדל מובהק (F3,156=3.42, p<0.05; איור 2). גדורות ללא רעייה הראו עלייה מובהקת בביומסה מפברואר לאפריל (F1,32=21.65, p<0.001). לעומת זאת, טיפולי עוצמת הרעייה הנמוכה, הבינונית והגבוהה לא הראו שינוי מובהק בביומסה (F1,32=0.53, p=0.47; F1,32=2.22, p=0.15; F1,32=0.14, p=0.71, בהתאמה; איור 2).
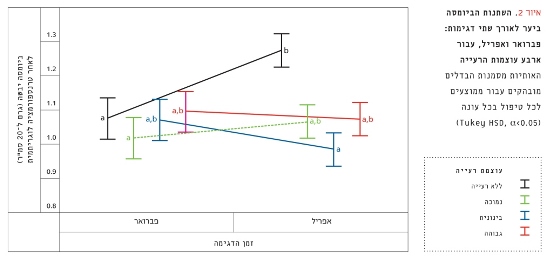
איור 2
השתנות הביומסה ביער לאורך שתי דגימות: פברואר ואפריל, עבור ארבע עוצמות הרעייה
האותיות מסמנות הבדלים מובהקים עבור ממוצעים לכל טיפול בכל עונה (Tukey HSD, α<0.05)
מגוון מינים
מגוון המינים השתנה באופן מובהק עם טיפולי הרעייה ( F3,106=4.66, p<0.01; איור 3). עוצמת הרעייה הבינונית הובילה לעלייה הגדולה ביותר במגוון המינים (F1,16=31.88, p<0.001). עוצמת הרעייה הנמוכה הראתה עלייה קטנה יותר במגוון המינים (F1,23=23.41, p<0.001), ועוצמת הרעייה הגבוהה הראתה עלייה קטנה עוד יותר (F1,18=14.57, p<0.01). הגדורות ללא רעייה לא הראו שינוי מובהק במגוון המינים (F1,24=1.19, p=0.28).
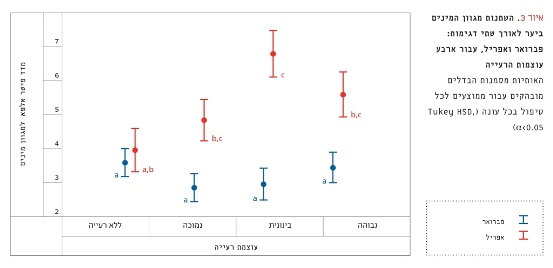
איור 3
השתנות מגוון המינים ביער לאורך שתי דגימות: פברואר ואפריל, עבור ארבע עוצמות הרעייה
האותיות מסמנות הבדלים מובהקים עבור ממוצעים לכל טיפול בכל עונה (Tukey HSD, α<0.05)
קבוצות תפקודיות והרכב מינים
פרופורציות הקבוצות תפקודיות לא הראו אינטראקציה מובהקת בין זמן הדגימה לעוצמת הרעייה (רחבי עלים F3,156=0.83, p=0.48; דגניים נמוכים F3,156=1.3, p=0.28; דגניים גבוהים F3,156=0.45, p=0.71; קטניות F3,156=2.01, p=0.11; קוצניים F3,156=0.53, p=0.66). למרות זאת, כמה קבוצות כן הראו הבדלים מובהקים תחת עוצמות רעייה שונות: הקטניות ירדו בפרופורציה מפברואר לאפריל בחלקות ללא רעייה (F1,32=8.99, p<0.05), ודגניים גבוהים ורחבי עלים ירדו בפרופורציה מפברואר לאפריל תחת עוצמת רעייה גבוהה (F1,32=8.32, p<0.05, F1,32=9.55, p<0.01 בהתאמה). באפריל הראו הדגניים הנמוכים פרופורציה גבוהה יותר בעוצמת הרעייה הגבוהה ביחס לחלקות ללא רעייה (F1,78=5.30, p<0.05).
ניתוח nMDS הראה הבדל בהרכב המינים לפני רעייה בעוצמה גבוהה ואחריה (Stress=0.18; איור 4). בגדורות ללא הרעייה, לעומת זאת, לא ניכר הבדל גדול בהרכב המינים לפני הרעייה ואחריה (Stress=0.23; איור 4).
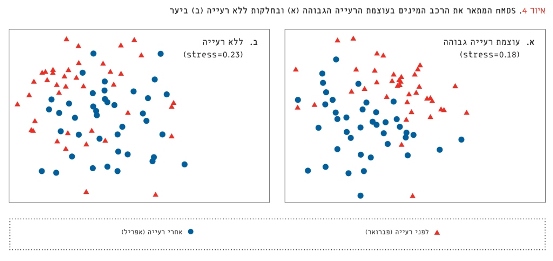
איור 4
nMDS המתאר את הרכב המינים בעוצמת הרעייה הגבוהה (א) ובחלקות ללא רעייה (ב) ביער
דיון ומסקנות
המטרה העיקרית של המחקר הייתה להבין כיצד עוצמות רעייה שונות משפיעות על ביומסה ועל מגוון מינים עשבוניים ביער איקליפטוס נטוע. באופן כללי, מצאנו במחקר כי רעיית צאן מנעה עלייה עונתית בביומסה, אך הובילה לשינויים במגוון ובהרכב של המינים העשבוניים: רעייה גרמה לעלייה במגוון המינים, והעלייה הגדולה ביותר נמדדה לאחר רעייה בעוצמה בינונית. חשיבות המחקר נובעת מכך שזהו המחקר הראשון שבוחן השפעות קצרות טווח של רעייה על עשבייה ביער איקליפטוס בצפון-מערב הנגב. תוצאות המחקר יכולות לשמש פרוטוקול עבור יערנים ומנהלי שטחים פתוחים המעוניינים לשלב רעיית צאן בשטחם ביעילות ובניהול נכון.
בחלקות שלא הייתה בהן רעייה, הביומסה עלתה במשך הניסוי. תוצאה זו דומה לתוצאות בניסויים דומים שנמנעה בהם רעייה באזורים יובשניים [14, 15], והיא מראה כי בהתאם להשערות, מניעת רעייה מובילה לעלייה בביומסה, דבר שמוביל להצטברות חומר דליק. לעומת זאת, בחלקות שנחשפו לעוצמות רעייה שונות, הביומסה לא הראתה עלייה או ירידה במהלך העונה. עם זאת, ניתן לראות כי הביומסה הראתה מגמת ירידה בעוצמת הרעייה הבינונית. ייתכן שבשל הסגירה והפתיחה של הגדורות, הצמחייה בגדורת העוצמה הבינונית הספיקה להתאושש במהלך היותה סגורה, והצאן נמשך יותר לצמחייה בה.
מגוון המינים הראה דפוס הדומה לאונימודלי (התפלגות בעלת נקודת מקסימום אחת), ועוצמת הרעייה הבינונית הניבה את העלייה הגבוהה ביותר במגוון המינים. בגדורות ללא רעייה מגוון המינים לא השתנה במהלך עונת הרעייה. מתוצאות אלה ניתן להבין כי מומלץ לשלב רעייה ביערות כדי להגדיל את מגוון מיני העשבייה.
עוצמת הרעייה הבינונית גרמה למגוון המינים הגבוה ביותר וכן לאי-שינוי בביומסה העשבונית. אם כך, ניתן להסיק כי עוצמות רעייה בינוניות הן הנדרשות כדי לענות על הצרכים של מנהלי הממשק של היער. אי-רעייה גורמת להצטברות ביומסה דליקה ולמגוון מינים נמוך. לעומת זאת, רעייה בעוצמה גבוהה מדי עלולה להוריד גם היא את מגוון המינים העשבוניים, דבר שיפגע בתפקוד האקולוגי ובתיירות.
הקבוצות התפקודיות של הצמחים לא הראו מגמה ברורה של השתנות בעוצמות רעייה שונות. ייתכן שהסיבה לכך היא הטווח הקצר של המחקר. במחקרי רעייה שנמצא בהם שינוי בפרופורציות של קבוצות תפקודיות, נעשו ניסויים ארוכי טווח בני כמה שנים בכרי מרעה בצפון הארץ [12, 23]. עם זאת, הדגניים הנמוכים אכן הראו כיסוי גדול יותר בחלקות עם רעייה חזקה מאשר בחלקות ללא רעייה, דבר שמעיד על הפקת תועלת מסביבה מופרעת.
לסיכום, ניתן להמליץ על רעיית צאן ביערות האיקליפטוס הנטועים בצפון-מערב הנגב, שכן הרעייה מונעת הצטברות ביומסה דליקה בתת-היער וכן משפרת את מגוון מיני העשבייה. על הרעייה לא להיות בעוצמה גבוהה מדי, ויש לדאוג לפיזורה באופן הומוגני במרחב, כך שתתקבל עוצמת רעייה בינונית בכל רצפת היער. מכיוון שמחקר זה הוא ראשוני ובוצע לאורך עונה אחת בלבד, אנו ממליצים להמשיך ולבחון את השפעות הרעייה על עשביית יערות נטועים בטווח הארוך כדי להבין טוב יותר את הדינמיקה שבין הרעייה לבין הביומסה ומגוון המינים העשבוני.

תודות
ברצוננו להודות לשמשון שוקר, לטניה גנדלר, לזהבה סיגל וללירון סאמרפילד על סיוע בעבודת השטח ובזיהוי הצמחים. כמו כן אנו מודים לעופר עובדיה על ייעוץ סטטיסטי, ליערני קק"ל בנחל אסף ולרועים עודה ומוסא על שיתוף הפעולה.
- לממשק רעיית צאן מיוחסות השפעות חיוביות על המערכת הצמחית, כגון הפחתת סכנת השרפות והגדלת המגוון הביולוגי.
- על אף הידע הרב בנושא השפעות רעייה על מערכות צומח אחרות )עשבוניות וחורש ים תיכוני( יש פער ידע באשר להשפעות רעייה בשטחים שניטעו בהם עצי איקליפטוס.
- שכיחותם של יערות איקליפטוס נטועים בצפון- מערב הנגב מצריכה גיבוש גישה שתאפשר קיום מגוון מינים גבוה, אך גם תמנע התפרצות שרפות והתפשטותן.
- המאמר מספק בסיס ראשוני לקביעת מדיניות לשילוב רעיית עדרים במערכות צומח ביער איקליפטוס נטוע, כולל קביעת משטר הרעייה המיטבי ביער, שישלב מניעת שרפות עם קיום מרבי של מגוון מיני צומח.
המערכת
מקורות
- זליגמן נ, אונגר י, הנקין ז ואחרים. 2016. על צומח, בעלי חיים ואנשים: תורת ניהול המרעה בישראל. ירושלים: נקודת ח"ן.
- Asner GP, Elmore AJ, Olander LP, et al. 2004. Grazing systems, ecosystem responses, and global change. Annual Review of Environment and Resources 29: 261-299.
- Connell JH. 1978. Diversity in tropical rain forests and coral reefs. Science 199: 1302-1310.
- Diaz S, Lavorel S, McIntyre S, et al. 2007. Plant trait responses to grazing – A global synthesis. Global Change Biology 13: 313-341.
- Evlagon D, Kommisarchik S, Gurevich B, et al. 2012. Estimating normative grazing capacity of planted Mediterranean forests in a fire-prone environment. Agriculture, Ecosystems & Environment 155: 133-141.
- Fisher RA, Corbet AS, and Williams CB. 1943. The relation between the number of species and the number of individuals in a random sample of an animal population. The Journal of Animal Ecology 12: 42-58.
- Hadar L, Noy-Meir I, and Perevolotsky A. 1999. The effect of shrub clearing and grazing on the composition of a Mediterranean plant community: Functional groups versus species. Journal of Vegetation Science 10: 673-682.
- Huston M. 1979. A general hypothesis of species diversity. The American Naturalist 113: 81-101.
- Milchunas DG, Sala OE, and Lauenroth W. 1988. A generalized model of the effects of grazing by large herbivores on grassland community structure. The American Naturalist 132: 87-106.
- Ne’eman G and Perevolotsky A. 2000. The management of burned forests in Israel. In: Ne’eman G and Trabaud L (Eds). Ecology, biogeography and management of Pinus halepensis and P. brutia Mediterranean pine forest ecosystems. Leiden: Buckhuys Publishers.
- Noy-Meir I and Seligman NG. 1979. Management of semi-arid ecosystems in Israel. In: Walker BH (Ed). Management of semi-arid ecosystems. Amsterdam: Elsevier Science.
- Noy-Meir I, Gutman M, and Kaplan Y. 1989. Responses of Mediterranean grassland plants to grazing and protection. Journal of Ecology 77: 290-310.
- Noy-Meir I. 1995. Interactive effects of fire and grazing on structure and diversity of Mediterranean grasslands. Journal of Vegetation Science 6: 701-710.
- Oba G, Vetaas OR, and Stenseth NC. 2001. Relationships between biomass and plant species richness in arid zone grazing lands. Journal of Applied Ecology 38: 836-845.
- Osem Y, Perevolotsky A, and Kigel J. 2002. Grazing effect on diversity of annual plant communities in a semi-arid rangeland: Interactions with small-scale spatial and temporal variation in primary productivity. Journal of Ecology 90: 936-946.
- Papanikolaou AD, Fyllas NM, Mazaris AD, et al. 2011. Grazing effects on plant functional group diversity in Mediterranean shrublands. Biodiversity and Conservation 20: 2831-2843.
- Peco B, Sánchez AM, and Azcárate FM. 2006. Abandonment in grazing systems: Consequences for vegetation and soil. Agriculture, Ecosystems & Environment 113: 284-294.
- Peco B, Carmona CP, de Pablos I, et al. 2012. Effects of grazing abandonment on functional and taxonomic diversity of Mediterranean grasslands. Agriculture, Ecosystems and Environment 152: 27-32.
- Perevolotsky A and Seligman NG. 1998. Role of grazing in Mediterranean rangeland ecosystems. Bioscience 48: 1007-1017.
- Proulx M and Mazumder A. 1998. Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems. Ecology 79: 2581-2592.
- Rosenzweig ML and Abramsky Z. 1993. How are diversity and productivity related? In: Ricklefs RE and Schulter D. Species diversity in ecological communities. Chicago: University of Chicago Press.
- Sternberg M, Gutman M, Perevolotsky A, et al. 2000. Vegetation response to grazing management in a Mediterranean herbaceous community: A functional group approach. Journal of Applied Ecology 37: 224-237.
- Sternberg M, Golodets C, Gutman M, et al. 2015. Testing the limits of resistance: A 19 yr study of Mediterranean grassland response to grazing regimes. Global Change Biology 21: 1939-1950.
- Watkinson AR and Ormerod SJ. 2001. Grasslands, grazing and biodiversity: Editors’ introduction. Journal of Applied Ecology 38: 233-237.



 מהדורה מודפסת
מהדורה מודפסת








