
חזית המחקר
הכרת המגוון הגנטי של מיני האלונים הגדלים בארץ – בסיס להצלחה בנטיעות
גלינה שקלר המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקני
לאוניד קורול המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקני

אלון מצוי סמוך לקיבוץ צובה – מהגדולים ומהעתיקים בהרי ירושלים. העץ נשמר מאות שנים בזכות קדושתו | צילום: גיא אסייג, באדיבות ארכיון קק"ל
מאת
גבריאל שילר
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקניגלינה שקלר
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקנילאוניד קורול
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקנימאת
גבריאל שילר
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקניגלינה שקלר
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקנילאוניד קורול
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי - מרכז וולקניבתחום היערנות, שעוסקת במיני צמחים שלא עברו ברֵרה מכוונת לשם טיפוח תכונות מסוימות, ידוע כי יש מתאם בין מגוון גנטי גבוה בצמח הבודד לצמיחה טובה שלו ולבריאותו. מכיוון שהדרישה לשתילים איכותיים של מיני אלונים מקומיים למטרות נטיעה בשטחים הפתוחים, ביערות, בפארקים ובגינות בארץ מתעצמת בשנים האחרונות, חשוב להכיר את הגנטיקה של אוכלוסיות עצי האלון בארץ. רצוי לאסוף את חומר הריבוי (בלוטים) כדי לגדל שתילים איכותיים מאוכלוסיות בעלות מגוון גנטי גבוה יחסית במסגרת המין. כדי לספק את המידע נחקרים המגוון הגנטי (השונות הגנטית) הכללי של המין, וכן המגוון בין ובתוך אוכלוסיות רבות שלו, בכל אחד משלושת מיני האלון העיקריים: אלון התבור, אלון התולע ואלון מצוי.
לשם קביעת המגוון הגנטי נעשה שימוש בסמנים גנטיים מולקולריים מסוג RAPD, משמע Random Amplified Polymorphic DNA, שהיו מקובלים במחקרים מסוג זה בעת ביצוע המחקר. השיטה הופעלה על די-אן-איי שהופק מעלים בריאים שנקטפו מכ-50 עצים בכל אוכלוסייה (בית גידול). דגימות נאספו מ-16 אוכלוסיות של אלון התבור, מ-24 אוכלוסיות של אלון מצוי ומ-14 אוכלוסיות של אלון התולע.
התוצאות של ספירת התדירויות של הסמנים הגנטיים באוכלוסייה ובמין שימשו בסיס לחישובים סטטיסטיים בעזרת תכנַת מומחה ייעודית. התוצאות שהתקבלו הראו כי המגוון הגנטי (השונות הגנטית במין – Ht) בשלושת מיני האלונים הוא: אלון התבור=0.414, אלון התולע=0.398 ואלון מצוי=0.354. ההבדלים בתדירות של הסמנים הגנטיים השונים באלון התבור אפשרו לקבץ את אוכלוסיותיו לשלוש התקבצויות גאוגרפיות-אקלימיות. התוצאות שהתקבלו עבור אלון מצוי הראו כי למרות הבדלים קטנים ביותר במבנה הגנטי בין האוכלוסיות, ניתן לקבץ אותן על פי האזורים הגאוגרפיים-אקלימיים העיקריים במדינה. לעומת זאת, התוצאות של הבדיקה הסטטיסטית בנתונים של אלון התולע הראו כי לא ניתן לגבש התקבצויות גאוגרפיות-אקולוגיות במין זה; לכל אוכלוסייה מקומית המבנה הגנטי שלה.
על קצה המזלג
- המאמר עוסק בשונות הגנטית של אלונים, מיני עצים מרכזיים בנוף הצומח הטבעי בישראל, שהם חלק נכבד מהעצים הניטעים כיום ביערות ישראל ובשטחים הפתוחים.
- במיני עצים שלא עברו ברֵרה מכוונת לשם טיפוח תכונות מועדפות, מוכר מתאם בין מגוון גנטי גבוה בעץ הבודד לבין מהירות גדילתו ועמידותו המשופרת בפני מזיקים ומחלות.
- יש צורך בהכרת השונות הגנטית בין אוכלוסיות ואזורים אקו-גאוגרפיים שונים, עבור כל אחד מהמינים בנפרד, כדי שמדיניות נטיעת האלונים תושפע, בין השאר, מהמבנה הגנטי של אוכלוסיות עצי האלון בארץ.
- ניתוח המגוון הגנטי בקרב מיני האלונים, מאפשר להצביע על אוכלוסיות נבחרות בקרב כל מין, שמומלץ לאסוף מהן חומר ריבוי איכותי למטרת נטיעות בכל אזור גאוגרפי-אקולוגי.
המערכת
הלכה למעשה
מבוא
החבל הים תיכוני בישראל ובירדן הוא גבול התפוצה הדרומי של שלושת מיני האלונים: אלון מצוי (Quercus calliprinos), אלון התבור (Quercus ithaburensis) ואלון התולע (Quercus boissieri), שגדלים בחורשות וביערות רחבי העלים בארץ [24]. אלונים אלה הם עמוד השדרה של תצורת צומח זו.
בשנת 1926 פִּרסם הממשל הבריטי את פקודת היערות בארץ ישראל כדי לרסן את ההתנהלות ששלטה עד אז – ניצול בלתי מבוקר של צמחיית העצים בארץ. דבר זה אִפשר לצמחייה המעוצה להשתקם במידה מסוימת, בעיקר דרך גדילת נצרים רבים מהגדמים, אך גם מנביטת בלוטים וזרעים. הצמחייה שהשתקמה היא שריד ליערות שהאדם עשה בהם כרצונו מאז החל להתיישב באזורנו [3]. כלומר, האוכלוסיות של מינים אלה היו נתונות לברֵרה שהפעיל האדם כדי לספק את צרכיו הייחודיים במשך 10,000 שנה לפחות (50 דורות של 200 שנה – משך חיי העץ) – זמן מספיק כדי לגרום בוודאות שינויים במגוון הגנטי, על כל המשתמע מכך [10]. אך באותה מידה, אוכלוסיות אלה הן גם שריד לצמחייה שהתאימה את עצמה מבחינה גנטית ואקו-פיזיולוגית לתנאים שנוצרו בכל בית גידול בפני עצמו כתוצאה מהתדלדלות עומק הקרקע, מפוריותה וממשטר השרֵפות והרעייה בו. זה מכבר קיימת ההבנה שלתנאים האקולוגיים בכל בית גידול יש השפעה רבה על המגוון הגנטי של הפרטים ושל האוכלוסייה בכללותה אצל כל מין ומין, השפעה שהיא הבסיס להיווצרות אקוטיפים (אוכלוסיות מקומיות בתוך מין ביולוגי בעלות מערך התאמות גנטי מסוים לתנאי סביבה ייחודיים – ecotypes) במסגרת המינים. באותה מידה קיימת ההבנה שככל שהמגוון הגנטי בעץ הבודד גבוה יותר, כך גבוהה יותר עמידותו בפני מזיקים ומחלות מצד אחד, והתפתחותו טובה יותר מצד שני [ 5, 8, 9, 11, 12].
כל אחד משלושת מיני האלונים שגדל בארץ בתחום החבל הים תיכוני, גדל בבתי גידול שנבדלים זה מזה בתנאיהם האקולוגיים (המסלע, הקרקע, המיקרו-אקלים, כמות הגשמים ועוד). באחדים מהגורמים האקולוגיים בישראל קיים שינוי רציף מצפון לדרום וממערב למזרח (למשל, פחיתה בכמות הגשם הרב-שנתית הממוצעת). היות שכך, נוצר הצורך להרחיב את הידע על המגוון הגנטי בקרב האלונים בארץ, כדי שתתאפשר בחירה של האוכלוסייה (האקוטיפ) "הנכונה", ומתוכה – של העצים המתאימים מהבחינה הפנוטיפית (ביטוי חיצוני לתכונות גנטיות) לשם איסוף חומר ריבוי מהם (עצי האֵם לזרעים-בלוטים). כך תתאפשר היענות לדרישה הגוברת לשתילים איכותיים, שנובעת מעלייה בכמות נטיעת אלונים מקומיים ביערות, בשטחים הפתוחים ובגינון העירוני והפרטי כעצי נוי. כלומר, המטרה היא קבלת שתילים בעלי מגוון גנטי גבוה יחסית בכל אחד מהמינים, דבר שיאפשר להם להתמודד טוב יותר עם תנאי בית הגידול ולהתפתח בצורה טובה ובריאה לאורך שנים.
מתוך הצורך הזה, נערכו מספר מחקרים [17, 18, 19, 20], שמטרתם הייתה להגדיר בראשונה את המגוון הגנטי הכללי בתוך ובין האוכלוסיות במסגרת כל מין משלושת מיני האלון העיקריים בנוף הארץ: אלון מצוי, אלון התבור ואלון התולע. כמו כן, נבדק הקשר בין המגוון הגנטי לבין האזורים הגאוגרפיים-אקולוגיים במדינה. המאמר שלפניכם מסכם את סדרת המאמרים כדי להנחיל ידע זה לציבור הרחב.
חומרים ושיטות
קביעת האוכלוסיות שנכללו במסגרת המחקר נעשתה בהתייעצות עם מספר בוטנאים ואקולוגים, כדי שיתקיים ייצוג הולם לתפוצת כל מין במדינה ולכל אזור גאוגרפי-אקולוגי.
איסוף החומר הצמחי לשם הפקת הדי-אן-איי
לקביעת רמת השונות הגנטית בעצים בודדים, באוכלוסיות השונות ובמין בכללותו השתמשנו בדי-אן-איי שהופק מעלים בוגרים, שעל פי מראה עיניים לא היו נגועים במזיקים ובמחלות, ולא ניכרה בהם פגיעה מגורמים אחרים. האוכלוסיות (בתי גידול) שנבחרו היו 24 אוכלוסיות של אלון מצוי שמתוכן שתי אוכלוסיות מירדן, 16 אוכלוסיות של אלון התבור ו-14 אוכלוסיות של אלון התולע (ראו טבלה 1). מכל אחת מהן נדגמו 50 עצים (שהמרחק ביניהם היה גדול מ-25 מטר), ומכל אחד מהם נקטף חופן עלים. העלים שנקטפו מכל עץ נשמרו בנפרד במקרר עד להפקת הדי-אן-איי בשיטות המקובלות, שלא יפורטו כאן.
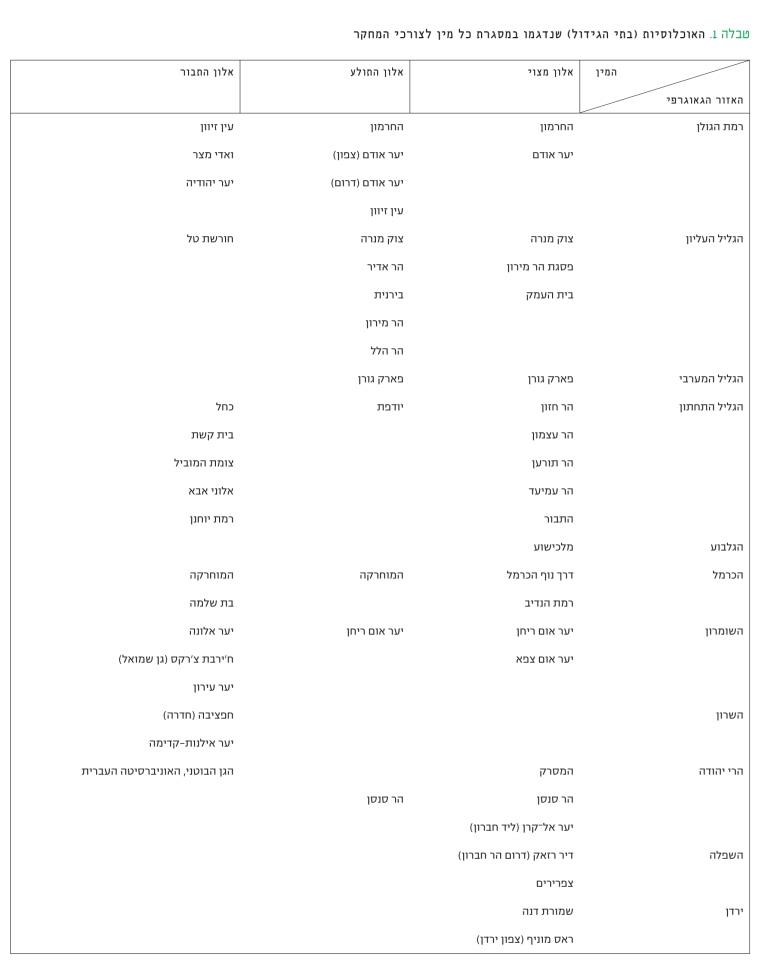
טבלה 1
האוכלוסיות (בתי הגידול) שנדגמו במסגרת כל מין לצורכי המחקר
אנליזה של הדי-אן-איי
האנליזה נעשתה בשיטת RAPD, משמע Randon Amplification of Polymorphic DNA, שהייתה מקובלת בסוף שנות ה-90 ובתחילת המאה ה-21 [4, 22], וזאת משתי סיבות עיקריות. סיבה אחת הייתה העלות הנמוכה, שאפשרה אנליזה של פרטים רבים. סיבה שנייה הייתה ששיטה זו לא הצריכה ידע גנטי מקדים, שכן רצף הגנום של האלון טרם פוענח, ומידע לגבי סמנים מולקולריים אחרים לא היה בנמצא. בשיטת RAPD משתמשים בתְחַלים (primers) קצרים יחסית לשם ריבוי מקטעי די-אן-איי אקראיים תוך שימוש בפי-סי-אר (PCR), משמע Polymerase Chain Reaction, והפרדתם בעזרת שדה חשמלי (electrophoresis) בג'ל אגרוס. מתוך 60 תחלים של חברת Operon שנבחנו לגבי מידת התאמתם, נעשה שימוש ב-20 שלאחר הרצת ביקורת בשדה חשמלי הניבו את התוצאות המהימנות והברורות ביותר לפענוח. 20 התחלים יצרו בסך הכול 127 מקטעי די-אן-איי (=סמנים גנטיים); כל תַּחַל בוטא באמצעות 2-10 מקטעי די-אן-איי בעלי גודל (משקל) מולקולרי שונה. בדי-אן-איי של אלון התבור זוהו 72 מקטעים פולימורפיים (polymorphic markers); בדי-אן-איי של אלון התולע 32 מקטעים; בדי-אן-איי של האלון המצוי התקבלו תוצאות מהימנות וברורות רק מ-10 תחלים, שיצרו 23 מקטעים. לאחר האלקטרופורֵזה צולמו הג'לים, ונעשתה ספירה של נוכחות מקטעי די-אן-איי בהתאם לתחלים לכל עץ בנפרד.
חישובים סטטיסטיים
החישובים נעשו בעזרת מגוון תכנוֹת סטטיסטיות ייעודיות שהיו קיימות בסוף שנות ה-90 ובתחילת המאה ה-21. המגוון הגנטי של המין בתוך כל אוכלוסייה (בית גידול) ובין האוכלוסיות חושב בעזרת תכנַת מומחה ייעודית: Popgene, version 1.32 [23]. כדי להעריך את השונות הגנטית בכל אוכלוסייה חושב מדד המגוון הגנטי של שאנון [13]. בכל אתר נבחן הגנוטיפ על פי העץ והתַחַל [14]. כמו כן, נבחנה רמת הבידול הגנטי בין קבוצות של אוכלוסיות [15]. המגוון הגנטי הכללי (Ht) חושב על סמך התדירות הממוצעת של כל מקטעי הדי-אן-איי שיצרו התחלים במסגרת המין (בכל העצים ובכל האוכלוסיות). המגוון הגנטי הממוצע בכל אוכלוסייה (Hs) חושב על סמך התדירות הממוצעת של כל מקטעי הדי-אן-איי בעצים בכל אוכלוסייה לחוד. הבידול בין אוכלוסיות (Gst) חושב בעזרת תכנַת Popgene כיחס Ht-Hs)/Ht). לשם השוואות מרובות של הבדלים בין אוכלוסיות או התקבצויות של אוכלוסיות (AMOVA) השתמשנו בתכנַת Sigmastat Statistical. המרחק הגנטי בין אוכלוסיות במסגרת המין [16] חושב תוך התחשבות בכל אתר. היחסים בין מרחקים גאוגרפיים וגנטיים בין האוכלוסיות במסגרת כל מין נקבעו בעזרת מבחן מאנטל תוך שימוש בתכנַת Arlequin 2.0 [21]. בוצע מבחן התקבצויות (cluster analyses) שנעשה בשיטת UPGMA, הכלולה בתכנָה PHYLIP 3.5 [6, 7].
תוצאות
תוצאות החישובים הסטטיסטיים לכל אוכלוסייה בתוך המין מסוכמות בנספח 1.
אלון התבור
המגוון הגנטי הכללי (Ht), שחושב על סמך התדירות הממוצעת של כל מקטע די-אן-איי מבין 72 המקטעים שיצרו 20 התחלים, הוא 0.414. עיון בתוצאות שבנספח 1 מראה שממוצע המגוון הגנטי בקרב האוכלוסיות שנדגמו (Hs) הוא 0.020 ± 0.362. האוכלוסייה בעלת המגוון הגנטי הגבוה ביותר (0.396) היא אוכלוסיית צומת המוביל, וזו עם המגוון הנמוך ביותר (0.323) היא חורשת טל. ממוצע (Gst), שהוא החלק מהמגוון הגנטי של המין שנובע מהבדלים בין האוכלוסיות במגוון הגנטי שבהן (Hs), הוא 0.046 ± 0.125. כלומר, 12.5% מהמגוון הגנטי הכללי מקורו בהבדלים בין האוכלוסיות. רגרסיה לינֵארית מובהקת נמצאה בין התדירות של מקטעי די-אן-איי שיצרו שני תחלים שונים לבין הרוחב הגאוגרפי של מיקום האוכלוסייה על פי רשת ישראל.
- רוחב גאוגרפי = (תדירות מקטעי התחל 1127.615XOPA05/2) מינוס 2771.335. R=0.596, p=0.015.
- רוחב גאוגרפי = (תדירות מקטעי התחל 1273.780XOPA12/2) פלוס 1377.952. R=0.529, p=0.035. רגרסיה לינֵארית נוספת, על גבול המובהקות, נמצאה בין התדירות של סמן גנטי מסוים לבין רמת המשקעים בבית הגידול.
- משקעים = (תדירות מקטעי התחל 528.283XOPA01/2) מינוס 1028.176. .R=0.453, p=0.078. גם בין המרחק הגאוגרפי למרחק הגנטי בין האוכלוסיות נמצאה רגרסיה לינֵארית, והמרחק הגאוגרפי הממוצע בין כל הזוגות האפשריים של אוכלוסיות אלון התבור שנבדקו (n=208) הוא 30.0 ± 47.5 ק"מ (הוא נע בין 5 ל-145 ק"מ).
- מרחק גנטי = 0.067343X מרחק גאוגרפי [ק"מ]. R=0.259, p=0.005, n=208.
מבחן ההתקבצויות הראה כי קיים הבדל מובהק בתדירות מקטעי הדי-אן-איי שיצרו שלושה מבין עשרים התחלים שהשתמשנו בהם, ונמצאה התאמה לאזורים גאוגרפיים שונים במדינה. תוצאה זו מאפשרת לקבץ את האוכלוסיות השונות לשלוש קבוצות (ראו טבלה 2): קבוצת הגולן והגליל העליון שכוללת את האוכלוסיות הגדלות בעין זיוון, בחורשת טל, בוואדי מצר וביער יהודיה; קבוצת הגליל התחתון שכוללת את האוכלוסיות שגדלות באלוני אבא, בבית קשת, בצומת המוביל ובח'ירבת צ'רקס; קבוצת הכרמל, השומרון והשרון שכוללת את האוכלוסיות שגדלות באילנות-קדימה, מצפון לחדרה, ביער עירון, בבת שלמה, במוחרקה וברמת יוחנן. האוכלוסיות כחל ויער אלונה קובצו ביחד כקבוצה נפרדת למרות המרחק הגאוגרפי ביניהן.
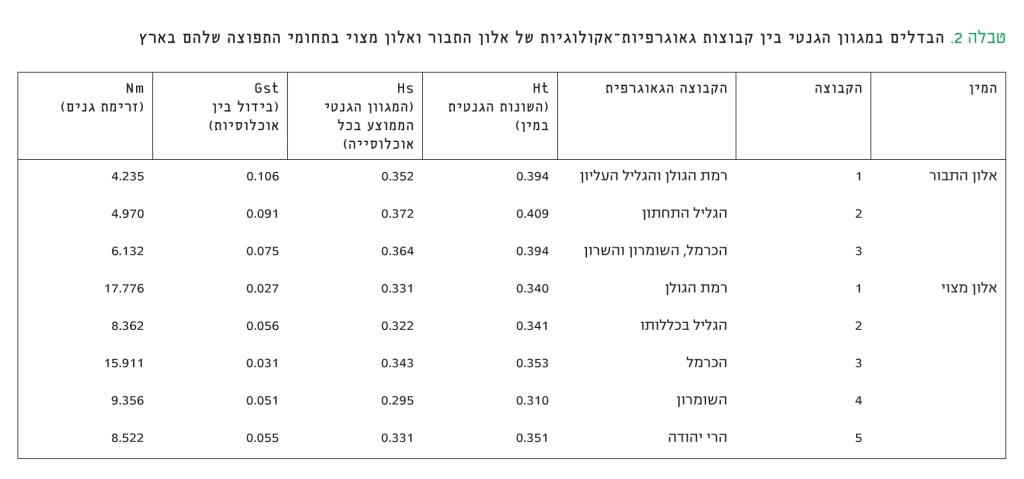
טבלה 2
הבדלים במגוון הגנטי בין קבוצות גאוגרפיות-אקולוגיות של אלון התבור ואלון מצוי בתחומי התפוצה שלהם בארץ
אלון התולע
חישובי המגוון הגנטי הכללי (Ht) במין זה העלו שהוא 0.398; מעיון בנספח 1 עולה כי ממוצע המגוון הגנטי בקרב האוכלוסיות שנדגמו (Hs) הוא 0.023±0.335; האוכלוסייה שהמגוון בה הוא הגבוה ביותר (0.369) גדלה במוחרקה בכרמל, וזו בעלת המגוון הנמוך ביותר (0.292) גדלה בהר אדיר. ממוצע (Gst) שמקורו בהבדלים שבין האוכלוסיות היה 0.057±0.162, כלומר 16.2% מהשונות הכללית. האפשרות שניתן יהיה לקבץ, בהתאם לתנאים הגאוגרפיים-אקלימיים שבמדינה, את האוכלוסיות של אלון התולע שנחקרו (טבלה 1), נבדקה בעזרת ניתוח סטטיסטי של ההתפלגות בתדירות של הסמנים הגנטיים (AMOVA). לא נמצא קשר סטטיסטי כלשהו בין תוצאות הבדיקות הגנטיות ובין המיקום הגאוגרפי של כל אוכלוסייה ואוכלוסייה.
אלון מצוי
חישוב המגוון הגנטי הכללי (Ht) במין זה העלה שהוא 0.354. מהתוצאות שבנספח 1 עולה כי ממוצע השונות הגנטית בקרב האוכלוסיות שנדגמו (Hs) הוא 0.016 ± 0.325. האוכלוסיות שהמגוון הגנטי בהן הוא הגבוה ביותר הן יער אודם (0.340), רמת הנדיב (0.348), דיר רזאק (0.355) וצפרירים (0.341). האוכלוסייה שהמגוון הגנטי בה הוא הנמוך ביותר היא יער אום-ריחן (0.288); רק 0.043 ± 0.119 מהשונות הכללית נובע מהבדלים בין האוכלוסיות (Gst). נעשה שימוש ב-AMOVA כדי לבדוק את ההתפלגות של תדירות מקטעי די-אן-איי בין כל אחת מחמש הקבוצות הגאוגרפיות-אקולוגיות – הגולן, הגליל, הכרמל, השומרון, הרי יהודה והשפלה הפנימית. מבדיקת ההתפלגות בין ובתוך (עצים בודדים) האוכלוסיות הנכללות בכל קבוצה, נמצא שמרב השונות הגנטית של המין – 99.97% – נמצאת בתוך האוכלוסיות; ההבדל הגנטי בין קבוצות גאוגרפיות-אקולוגיות הוא רק 0.13%, אך מובהקות ההבדלים היא P<0.0001. ההבדל הגנטי בין האוכלוסיות המרכיבות כל אחת מהקבוצות הוא 0.8%, ומובהקות ההבדלים היא P<0.0001. נוסף על כך, חושב ההבדל בין זוגות של אוכלוסיות שנובע משוני במבנה הגנטי (Fst). נמצא כי ב-13 זוגות ההבדל הגנטי בין אוכלוסיות היה מובהק ברמה של P<0.05 וללא קשר למרחק הגאוגרפי ביניהם. רק שתי אוכלוסיות, צפרירים ודיר רזאק, שגדלות באדמת רנדזינה בקצה הדרומי של תחום התפוצה הטבעי של האלון המצוי בארץ, הראו מרחק גנטי רב מכל שאר האוכלוסיות שנבדקו. נמצא כי האוכלוסייה של פארק גורן בגליל המערבי נבדלת גנטית מ-20 מתוך 24 האוכלוסיות שנבדקו. לעומת זאת, ערכי ה-Fst של אוכלוסיות בית העמק, פארק השרון והר עמיעד היו דומים ל-15 אוכלוסיות אחרות במדינה, שכל אחת מהן גדלה בתנאים אקולוגיים שונים. את ההבדלים בין הקבוצות במדדים גנטיים אפשר לראות בטבלה 3. ההבדל הבולט לעין הוא בזרימת גנים (Nm) בתוך הקבוצות השונות. זרימת הגנים ברמת הגולן ובכרמל רבה מאוד וכמעט כפולה מזו שבשאר הקבוצות.
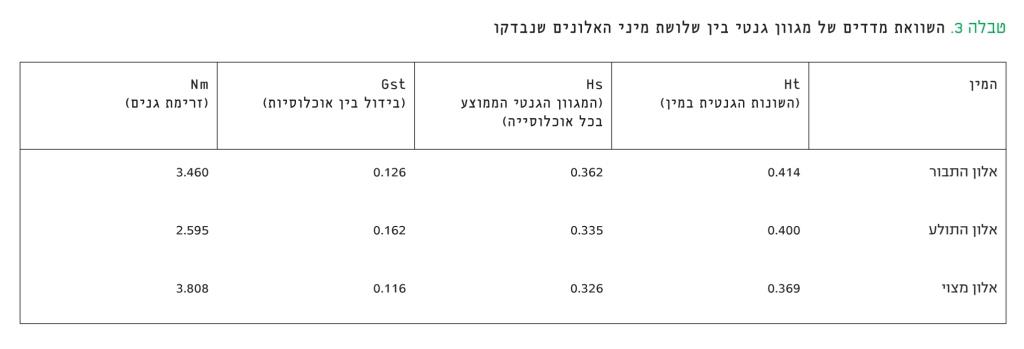
טבלה 3
השוואת מדדים של מגוון גנטי בין שלושת מיני האלונים שנבדקו
דיון
המחקר הגנטי הראשוני הזה, שנעשה במיני האלונים בארץ לפני כעשור, הוא המחקר היחידי עד כה המאפשר לסמן אוכלוסיות שהמגוון הגנטי בהן הוא הגדול ביותר במסגרת הקבוצות הגאוגרפיות-אקולוגיות במדינה. היות שידע יערני מעשי ומחקרי הראה כי עצים בעלי מגוון גנטי גבוה יחסית לזה של המין, גדלים טוב יותר ועמידים יותר למזיקים ולמחלות, רצוי לאסוף חומר ריבוי מאוכלוסיות אלה לשם יצירת שתילים איכותיים לנטיעה במסגרת האזורים השונים במדינה [4, 7, 8, 9, 10, 11].
להלן ההמלצות ביחס לאיסוף בלוטים לצורך ריבוי המינים השונים:
- ריבוי אלון התבור – רצוי לאסוף את הבלוטים מהאזורים הבאים: ברמת הגולן ובגליל העליון – מיער יהודיה ומכחל; באזור הגליל התחתון – מצפון-מזרח לצומת המוביל הארצי; באזור הכרמל, בשומרון ובשרון – מיער עירון.
- ריבוי האלון המצוי – למרות ההבדל הגנטי המזערי בין אוכלוסיות האלון המצוי בארץ רצוי מאוד לאסוף בלוטים לשם יצירת שתילים איכותיים מהאזורים הבאים: ברמת הגולן – מיער מסעדה; בגליל העליון – מיער בית העמק; בגליל התחתון – מיער הר תורען או מהתבור; בגלבוע – מיער מלכישוע; בשומרון, בהרי יהודה, בשפלה הפנימית ובדרום – מיער דיר רזאק או מצפרירים, שהמגוון הגנטי בהם גדול, והעצים בהם צומחים כבר עתה באזור צחיח.
- ריבוי אלון התולע – עץ זה אינו מתקבץ לקבוצות בהתאם לתנאים גאוגרפיים-אקלימיים. רצוי לאסוף בלוטים מאוכלוסיות שהמגוון הגנטי בהן הוא הגבוה ביותר – בחלק הדרומי של יער מסעדה, בחרמון, במוחרקה ובפארק גורן.
נוסף על כך, העוסקים בגידול אלונים צריכים להימנע מאיסוף בלוטים מן הקרקע, ולאסוף את הבלוטים רק מעצים שהפנוטיפ שלהם תואם את הקריטריונים של עץ האֵם לחומר ריבוי. אחד הקריטריונים הוא שיש לעץ שלטון בולט של קודקוד הצמיחה המרכזי [2]. אין לאסוף את הבלוטים מהקרקע (אף על פי שהדבר נוח יותר), מאחר ששם נוצרת תערובת מכל העצים השכנים.
במסגרת המחקר במגוון הגנטי באלון התבור, נמצא כי יש אוכלוסיות שגדלות באזור גאוגרפי מסוים, בעוד המגוון הגנטי בהן משייך אותן לאזור גאוגרפי אחר. לדוגמה, מסתבר כי חורשת טל שבעמק החולה, קשורה ליער יהודיה. דבר זה מרמז כי חומר גנטי שנאסף ביער יהודיה שימש לנטיעת החורשה (ואכן עד היום ניכרים בחורשה זו השורות). אוכלוסיית ח'ירבת צ'רקס שליד פרדס חנה קשורה לקבוצה של הגליל התחתון, וזאת למרות המרחק הגאוגרפי בין המקומות. ניתן להסביר ממצא זה בכך שמדובר באוכלוסייה נטועה, ושמקור הבלוטים בה היה באזור הגליל התחתון. כמו כן, אוכלוסיית כחל קשורה לקבוצת רמת הגולן והגליל העליון. אין לנו הסבר לתופעה זו, מעבר לכך שנראה כי היא נוסדה בעזרת נטיעת חומר ריבוי שנאסף ברמת הגולן. נראה שבעבר נהגו לנטוע את אלון התבור במטרה לספק בלוטים למאכל אדם ובעלי חיים [1].

סיכום
מאמר זה מסכם ממצאים ממחקר גנטי ראשוני ויחידי בקרב שלושה מיני אלונים הגדלים בחבל הים תיכוני בארץ. תוצאות המחקר מעידות על המגוון הגנטי בקרב מיני האלונים, ומציעות אוכלוסיות נבחרות בקרב כל מין ומין, שמהן רצוי לאסוף חומר ריבוי איכותי לנטיעות למטרות גינון ולנטיעות ביערות ובשטחים הפתוחים בכל אזור גאוגרפי-אקולוגי במדינה.
- המאמר עוסק בשונות הגנטית של אלונים, מיני עצים מרכזיים בנוף הצומח הטבעי בישראל, שהם חלק נכבד מהעצים הניטעים כיום ביערות ישראל ובשטחים הפתוחים.
- במיני עצים שלא עברו ברֵרה מכוונת לשם טיפוח תכונות מועדפות, מוכר מתאם בין מגוון גנטי גבוה בעץ הבודד לבין מהירות גדילתו ועמידותו המשופרת בפני מזיקים ומחלות.
- יש צורך בהכרת השונות הגנטית בין אוכלוסיות ואזורים אקו-גאוגרפיים שונים, עבור כל אחד מהמינים בנפרד, כדי שמדיניות נטיעת האלונים תושפע, בין השאר, מהמבנה הגנטי של אוכלוסיות עצי האלון בארץ.
- ניתוח המגוון הגנטי בקרב מיני האלונים, מאפשר להצביע על אוכלוסיות נבחרות בקרב כל מין, שמומלץ לאסוף מהן חומר ריבוי איכותי למטרת נטיעות בכל אזור גאוגרפי-אקולוגי.
המערכת
נספחים
נספח 1
להורדהמקורות
- אליאב א. 1985. שימוש בבלוטי אלון התבור למאכל. רתם (כתב עת לנושאי שדה בוטניים בארץ ישראל) 14: 73-72.
- טפר א. 1997. הגורמים המשפיעים על עיצוב צורת הגידול של האלון המצוי (עבודת גמר לתואר מוסמך). ירושלים: האוניברסיטה העברית בירושלים.
- פרבולוצקי א. 2013. האדם כמהנדס–על אקולוגי בחורש הים–תיכוני. בתוך: פרבולוצקי א (עורך). ממשק ושימור האקוסיסטמה הים תיכונית: רמת הנדיב כמשל. זכרון יעקב: רמת הנדיב.
- Bodenes D, Joandet S, Laigret F, and Kremer A. 1997. Detection of genomic regions differentiating two closely related oak species Quercus petraea (Matt.) Liebl. and Quercus robur L. Heredity 78: 433–444.
- Bush RM, Smouse PE, and Ledig FT. 1987. The fitness consequences of multiple-locus heterozygosity: The relationship between heterozygosity and growth rate in pitch pine (Pinus rigida Mill.). Evolution 41: 787–798.
- Cavalli-Sforza LL and Edwards AWF. 1967. Phylogenetic analysis: Models and estimation procedures. Evolution 21: 550–570.
- Felsenstein J. 1993. PHYLIP (Phylogeny Inference Package) Ver 3.5c. Department of Genetics, University of Washington, Seattle. WA. Focus 12: 13–15.
- Knowles P and Grant MT. 1981. Genetic patterns associated with growth variability in Ponderosa pine. American Journal of Botany 68: 942–946.
- Knowles P and Mitton JB. 1980. Genetic heterozygosity and radial growth variability in Pinus contorta. Silvae Gentica 29: 114–118.
- Ledig TF. 1992. Human impact on genetic diversity in forest ecosystems. Oikos 63: 87–108.
- Ledig FT, Guris RP, and Bonefeld B. 1983. The relation of growth to heterozygosity in pitch pine. Evolution 37: 1227–1238.
- Ledig FT and Kitzmiller JH. 1992. Genetic strategies for afforestation in the face of global climate change. Forest Ecology and Management 50: 153–169.
- Lewontin RC. 1972. The apportionments of human diversity. Evolution Biology 6: 381–398.
- Nei M. 1973. Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences U.S.A. 70: 3321–3323.
- Nei M. 1978. Molecular evolutionary genetics. New York: Columbia University Press.
- Nei M. 1987. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89: 283–292.
- Schiller G, Shklar G, and Korol L. 2003. Genetic diversity assessment by random amplified polymorphic DNA of oaks in Israel. 1. Tabor oak (Quercus aegilops L. ssp. ithaburensis [Decne] Boiss.). Israel Journal of Plant Sciences 51: 1–10.
- Schiller G, Shklar G, and Korol L. 2004. Genetic diversity assessment by random amplified polymorphic DNA of oaks in Israel. 2. Quercus boissieri Reut. Israel Journal of Plant Sciences 52: 315–322.
- Schiller G, Herr N, Shklar G, and Korol L. 2005. Diversity assessment, by means of random amplified DNA, of leaf phenology variation in Quercus ithaburensis Decne. Israel Journal of Plant Sciences 53: 75–78.
- Schiller G, Shklar G, and Korol L. 2006. Genetic diversity assessment by random amplified polymorphic DNA of oaks. 3. Quercus calliprinos Webb. in Israel and Jordan. Israel Journal of Plant Sciences 54: 137–148.
- Schneider S, Kueffer JM, Roessli D, and Excoffier I. 1997. ARLEQUIN, ver 1.1: Software for population genetic data analysis. Genetics and Biometry Laboratory, University of Geneva, Switzerland.
- Scotti-Saintagne C, Mariette S, Porth I, et al. 2004. Genome scanning for interspecific differentiation between two closely related oak species [Quercus robur L. and Q. petraea (Matt) Liebl.]. Genetics 168(3): 1615–1626.
- Yeh FC, Yang R-C, Boyle TBJ, et al. 1997. POPGENE ver 1.32. The user-friendly software for population genetic analysis. Molecular Biology and Biotechnology Center, University of Alberta, Canada.
- Zohary M. 1973. Geobotanical foundation of the Middle East. Stuttgart: Gustav Fisher Verlag.


 מהדורה מודפסת
מהדורה מודפסת








