
חזית המחקר
האם לאוכלי צמחים ישנן העדפות בין פרטים שכנים של אותו מין שיח בחורש? המקרה של העז ואלת המסטיק
חיים קיגל המכון למדעי הצמח, הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושלים
צח אהרון גלסר פארק הטבע רמת הנדיב
נתיב דודאי היחידה לצמחי תבלין, מרפא ובושם, מרכז מחקר נווה יער, מִנהל המחקר החקלאי – מרכז וולקני ([email protected])
יוג'ין דוד אונגר המחלקה למשאבי טבע, המכון למדעי הצמח ומרכז מחקר גילת, מִנהל המחקר החקלאי – מרכז וולקני

המחקר נערך בפארק הטבע רמת הנדיב. ברמת הנדיב עדר של 180 עיזים המשמש כלי ממשק לטיפול בחורש לצורך הגדלת מגוון בתי הגידול, הקטנת כיסוי הצומח המעוצה ומניעת שרפות. העדר רועה בפארק הטבע על פי תוכנית רעייה המותאמת לפארק. הרעייה בפארק מלווה בניטור השינויים בכיסוי הצמחי באזורי הרעייה השונים וגם במעקב אחר ביצועי העדר ואיכות החלב המופק לאחר ההזנה במרעה | צילום: ינינה זסלבסקי אפק
מאת
שילה נבון
המחלקה למשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי – מרכז וולקני; המכון למדעי הצמח, הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםחיים קיגל
המכון למדעי הצמח, הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםצח אהרון גלסר
פארק הטבע רמת הנדיבנתיב דודאי
היחידה לצמחי תבלין, מרפא ובושם, מרכז מחקר נווה יער, מִנהל המחקר החקלאי – מרכז וולקנייוג'ין דוד אונגר
המחלקה למשאבי טבע, המכון למדעי הצמח ומרכז מחקר גילת, מִנהל המחקר החקלאי – מרכז וולקנימאת
שילה נבון
המחלקה למשאבי טבע, המכון למדעי הצמח, מִנהל המחקר החקלאי – מרכז וולקני; המכון למדעי הצמח, הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםחיים קיגל
המכון למדעי הצמח, הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםצח אהרון גלסר
פארק הטבע רמת הנדיבנתיב דודאי
היחידה לצמחי תבלין, מרפא ובושם, מרכז מחקר נווה יער, מִנהל המחקר החקלאי – מרכז וולקנייוג'ין דוד אונגר
המחלקה למשאבי טבע, המכון למדעי הצמח ומרכז מחקר גילת, מִנהל המחקר החקלאי – מרכז וולקנירעיית עדרי עיזים בחורש ים תיכוני משמשת כלי לניהול שטחי החורש. השפעת הרעייה על בית הגידול תלויה ביחסי הגומלין בין בעלי החיים לצומח, נושא שנחקר בדרך כלל ברזולוציה שעד לרמת המין. מטרתו של מחקר זה היא לבחון את יחסי הגומלין הללו ברזולוציה מפורטת יותר, ולתת מענה לשאלה: האם עיזים במרעה, הניזונות מפרטים שכנים של מין משיחי החורש, בוחרות את מזונן באופן אקראי? כמין מודל למחקר נבחרה אלת המסטיק, משיחי החורש החשובים. במחקר קודם בחנו את פרופיל הרכיבים הנדיפים באוכלוסיית אלת המסטיק בחלקה מקומית בכרמל ששטחה כ-9 דונם (חלקת כבארה). נמצא כי השונות באוכלוסייה אינה רציפה, וכי 88% מהשיחים מתקבצים לשני טיפוסי נדיפים עיקריים: באחד בולטים germacrene D ו-limonene (להלן כימוטיפ L), ובאחר germacrene D ו-a-pinene (להלן כימוטיפ P). במחקר הנוכחי הורחב מיפוי השונות הכימית באוכלוסייה זו, ונבדקו מספר מדדים המשפיעים על האיכות התזונתית של השיחים: תכולת חלבון, טָנינים, סיבים ומינרלים ונעכלות העלווה. בוצעו מבחני העדפה בתנאים מבוקרים – לעיזים הוצגו זוגות ענפים משיחים שנדגמו מאוכלוסיית החלקה בהתאם להרכב הכימי. נמצאה העדפה מובהקת, בינונית בעוצמתה, מצד העיזים לשיחי אלת המסטיק מכימוטיפ L על פני אלה מכימוטיפ P. שלוש מתכונות השיח נמצאו בעלות קשר מובהק להעדפה בין שיחים: א. תכולת limonene (p=0.0002; R2=0.27) בהתאמה חיובית; ב. תכולת a-pinene (p=0.0015; R2=0.20) בהתאמה שלילית; ג. תכולת טָנינים (p<0.0001; R2=0.34) בהתאמה שלילית. ההתאמה הטובה ביותר התקבלה משילוב של שני הגורמים טָנינים ו-limonene (p<0.0001; R2=0.39). הממצאים מלמדים כי עיזים במרעה חורש ים תיכוני ערות להבדלים בין פרטים שכנים של אלת המסטיק, והדבר אף מתבטא בהתנהגות הרעייה שלהן. במאמר אנו מציעים מנגנון אפשרי המסביר את דפוס ההעדפה שנמצא אצל העיזים. ניתן להעריך כי העדפות בין צמחים שכנים מאותו המין קיימות גם אצל אוכלי צמחים נוספים הניזונים מאוכלוסיות של מינים מעוצים אחרים בחורש. לכן, בביצוע תצפיות על הנעשה בחורש, מומלצת ערנות גם למתרחש בתוך רמת המין.
על קצה המזלג
- לתחום המחקר של רעיית בעלי חיים צמחוניים בחורש ים תיכוני חשיבות רבה בגיבוש מדיניות ניהול שטחי החורש.
בחינת דפוסי שיחור המזון של עיזים משפרת את הבנת השפעתה של הרעייה על ממשק החורש. - המחקר מצא שיש שונות בין פרטים בני אותו מין באוכלוסיית הצומח (במקרה זה – אלת המסטיק) שגורמת להבדלים בשיעור צריכתם על–ידי עיזים. ממצא זה מחדד את ההבנה לגבי העדפות פרטניות של חיות מרעה.
- המאמר מוסיף נדבך להבנת היבט אקולוגי המשפיע על רעיית עיזים בחורש, שלה חשיבות במניעת שרפות ובשמירה על מגוון בתי גידול לחיות הבר ולמיני הצומח.
המערכת
הלכה למעשה
מבוא
החורש הוא תצורת צומח מרכזית בחבל הים תיכוני של ארץ ישראל. במשך אלפי השנים האחרונות חוותה צמחיית החורש פגיעה ניכרת, אך במהלך המאה האחרונה נחקקו חוקי הגנה על הצומח הטבעי, והחורש החל משתקם ומתחדש. בעקבות הירידה החדה בלחץ האדם והצאן החלו לחול שינויים במבנה החורש: עם חלוף השנים ניכר כי שיחי החורש משתלטים על בית הגידול וכך החורש נסגר, הנגישות אליו נפגעת, הצמחייה העשבונית נעלמת, ומגוון המינים בבית הגידול יורד בצורה חדה [6, 10]. החורש בארץ הוא מערכת מופרת, ובייחוד בולט בה היעדרם של אוכלי המעוצים (browsers) ובראשם אייל הכרמל (Capreolus capreolus) והיחמור הפרסי (Dama mesopotamica), שנכחדו מהארץ וכיום נעשים ניסיונות להשיבם. בעשורים האחרונים הובן כי אם ברצוננו לשמר את מגוון המינים המוכר ולמנוע השתלטות מלאה של שיחיית החורש על השטח, יש למצוא את הדרך לקיים בו הפרעה אקולוגית מתונה וקבועה המתמקדת בצומח המעוצה [34]. רעייה מבוקרת של עדרים בחורש היא כלי ישים וזול, המאפשר לשלוט בעיתוי ההפרעה ובעוצמתה, אך החורש הוא סביבת מרעה לא קלה לבעלי חיים אוכלי הצמחים (herbivores) [1, 4]. מבין חיות המרעה, העיזים (Capra hircus) הן המתאימות ביותר לרעייה בחורש. כרבים מבני הסוג יעל (Capra), העיזים מותאמות לאכילת עלווה של מינים מעוצים ולשגשוג בשטחי מרעה קשים, בעוד שההתאמה של פרות וכבשים לשטחים אלה פחותה [17, 32].
בנושא רעיית עדרי עיזים בחורש נערכו מחקרים מגוונים [למשל 20, 32], אך מרביתם בחנו את המערכת בסקאלה מרחבית רחבה וברזולוציה המגיעה לכל היותר עד לרמת מיני הצומח. עבודת מחקר זו מתמקדת באתרים מקומיים קטנים בחורש, ובאה לבחון את התנהגות הרעייה של העיזים ברזולוציה גבוהה, עד כדי התייחסות לרמת השיח הבודד. מטרתו הממוקדת של המחקר היא לתת מענה על השאלה: האם התפלגות הצריכה של עיזים במרעה הניזונות מאוכלוסיית שיחים שכנים מאותו המין תהיה בהכרח אקראית, או שישנן לעיזים העדפות בין פרטים?
למשרעת השונוּת הכימית בתוך אוכלוסיות מקומיות של שיחי החורש תפקיד מפתח בבירור שאלה זו [9]. עם זאת, מעטים המחקרים שבחנו באופן כמותי את השונות הגנטית והפנוטיפית בתוך אוכלוסיות של מיני המעוצים בחורש [5, 8, 22, 29, 31, 38]. יתר על כן, לא ברור אם המסקנות במחקרים אלה רלוונטיות גם עבור אוכלוסיות מקומיות הפרוסות על שטח מצומצם.
כמין מודל למחקר נבחרה אלת המסטיק (Pistacia lentiscus L.), משיחי החורש החשובים. לאלת המסטיק מקבץ רחב של רכיבים משניים ממגוון קבוצות כימיות: רכיבי שרף, רכיבי שמן אתרי וטָנינים (פוליפנולים המשקיעים חלבונים ופוגעים בערך התזונתי של המרעית) [23], ואף שאינה אהודה על פרות וכבשים, עיזים ניזונות ממנה, לעיתים בכמויות ניכרות [37]. תכולת הטָנינים הגבוהה נמצאה בעבר כמגבלה משמעותית לצריכה של עיזים משיח זה [12, 19]. לעומת זאת, השפעתם של רכיבי השרף והשמן האתרי שבאלת המסטיק על התנהגות הרעייה של עיזים טרם התבררה.
במחקר קודם [3] בחנו את פרופיל החומרים הנדיפים המרכיבים את השמן האתרי באוכלוסייה מקומית של אלת המסטיק בדרום הכרמל, בחלקה ששטחה 9.2 דונם (חלקת כבארה). הממצאים לימדו כי השונות הכימית באוכלוסייה איננה רציפה, וכי הרכבי הנדיפים מתקבצים לארבעה כימוטיפים (טיפוסים כימיים) מוגדרים היטב. 88% מהשיחים בחלקה השתייכו לאחד משני כימוטיפים: באחד בולטים germacrene D ו-limonene (להלן כימוטיפ L), ובאחר germacrene D ו-a-pinene (להלן כימוטיפ P). (48% ו-40% בהתאמה). אחת משאלות המפתח שעולות מהממצא היא אם לקיומם של טיפוסים כימיים שונים בבית הגידול המצומצם ישנן השלכות מעשיות על התנהגות שיחור המזון של העיזים הרועות. עקב ריבוי הגורמים הפוטנציאליים לצריכה שונה בין שיחים, כגון שיקולי תנועה ונגישות במרחב המצומצם, שאלה זו אינה יכולה להיבדק במלואה בשדה, אלא בניסוי מבוקר. עם זאת, ראינו חשיבות להדגים, גם אם בצורה מוגבלת, כי מקרים של העדפה בין שיחי אלת המסטיק שמקורם בתכונות השיחים עצמם, אכן מתרחשים בשדה ובתנאי אמת.
בתצפיות שדה מקדימות איתרנו בשטחי המרעה של פארק הטבע רמת הנדיב מספר זוגות שיחים שכנים של אלת המסטיק שניכרה בהם, לאחר שהעדר רעה בשטח, עדות ברורה להעדפה של העיזים את אחד השיחים בזוג על פני שכנו (איור 1). ניתוח כימי ראשוני מצא כי בהעדפות החדות שנצפו בשטח מעורבים הבדלים בפרופיל הנדיפים ובתכולתם הכללית [2]. תוצאות ראשוניות אלה, יחד עם השונות שמצאנו בהרכב הנדיפים באוכלוסיית אלת המסטיק שבחלקת כבארה [3], עוררו אותנו לבחון בתנאים מבוקרים את ההשפעה האפשרית של השונות הכימית הכוללת בחלקה מקומית זו, על התנהגות הרעייה של עיזים.
במחקר הנוכחי השלמנו את מיפוי השונות הכימית בחלקת כבארה על-ידי ביצוע בדיקות של מדדים תזונתיים בשיחים, וביצענו מבחני העדפה בתנאים מבוקרים: הצגנו לעיזים זוגות שיחים משני טיפוסי הנדיפים העיקריים. בוצעו שלושה טיפולים: LL – שני השיחים מכימוטיפ L; PP – שני השיחים מכימוטיפ P; LP – כל אחד מהשיחים מכימוטיפ שונה. הניתוחים הסטטיסטיים התבססו על מבחן t, על ANCOVA ועל רגרסיה ליניארית פשוטה ורב-גורמית. הכימוטיפים L ו-P אומנם מובחנים זה מזה בצורה חדה, אך ההבדל ביניהם אינו קיצוני. לכן, לא ידענו להעריך מראש אם תימצא העדפה ביניהם אם לאו. עם זאת, בהתבסס על המידע המוגבל המצוי בספרות [16, 36] השערתנו הייתה כי אם תימצא העדפה לאחד הכימוטיפים, הטיפוס L יועדף על פני הטיפוס P.
לפירוט פרק החומרים והשיטות המלא ראו נספח 1.

איור 1
עוצמת הרעייה השונה מדגימה בתנאי אמת את קיומה של העדפה אצל עיזים בין פרטים שכנים של אלת המסטיק
א-ד. ארבעה זוגות שונים של שיחים צמודים שנכללו בתצפית השדה המקדימה. העיזים הראו העדפה ברורה בצורה קיצונית לאחד מהשיחים;
ה. עז הניזונה מזוג השיחים המופיע גם בתמונה א. העז ניזונה משאריות השיח האהוד (מימין) אף על פי שכמעט לא נותרה עליו עלווה לאחר מספר ימי רעייה של העדר, ומתעלמת משכנו הדחוי למרות היותו עתיר–עלווה.
תוצאות
מדדים תזונתיים באוכלוסיית אלת המסטיק
להבדיל מהמבנה הבדיד של התפלגות הרכב הנדיפים באוכלוסיית אלת המסטיק שבחלקת כבארה [3], הערכים התזונתיים שנמדדו: חלבון (7.5-10.5%), טָנינים (19-25%), מינרלים (4.6-7.4%), ADL (13.5-16.5%), ADF (22.5-27.5%), NDF (30-36.5%) ונעכלות (31-39%), הראו כולם התפלגות ערכים רציפה על האוכלוסייה המקומית. לא נמצא כל הבדל בין הכימוטיפים L ו-P בערכי המדדים התזונתיים.
מבחני העדפה
משך כל פרק בדיקה בדיר היה 15:48±2:36 דקות. העז הקדישה לאכילה 88.5±8.4% מזמן פרק הבדיקה. ניתוח התוצאות התבסס על הבדלים בצריכת העלווה בזוגות השיחים שהוגשו לעיזים. צריכה יחסית משיח ימין (באחוזים) תסומן pRight, ומשיח שמאל pLeft, %pRight+pLeft=100.
העדפה בין שיחים מכימוטיפ L לשיחים מכימוטיפ P – ב-15 מתוך 18 החזרות שבוצעו בטיפול LP, צרכו העיזים מהשיח מטיפוס L יותר מאשר צרכו מהשיח מטיפוס P. בניתוח מבחן t בזוגות נמצאה העדפה מובהקת לטיפוס L על פני טיפוס P (p=0.0002, n=18, יחס צריכה ממוצע 2:1).
חשיבותם של הכימוטיפ והטָנינים באירועי העדפה בין שיחים – ערכי מדד ההעדפה pRight-pLeft שהתקבלו בטיפולים השונים לפי זהות הכימוטיפ היו: LL: -7.5±7.9%; PP: 14.1±7.0%; LP: -43±9.5%; PL: 24.3±10.0%. בניתוח השונות המשותפת (ANCOVA) נמצא כי הצירוף של זהות הכימוטיפים עם ההפרש בתכולת הטָנינים בין השיחים מסביר בצורה מובהקת (p<0.0001,R2=0.50) את ערכי מדד ההעדפה. שני הגורמים נמצאו בעלי תרומה מובהקת למודל (זהות הכימוטיפ: p=0.003; תכולת טָנינים: p=0.002).
הקשר בין נדיפים בודדים לבין העדפה – מבין 31 הרכיבים הנדיפים שנבחנו כגורמים משפיעים אפשריים על ההעדפה, נמצאו שני נדיפים כבעלי קשר ליניארי מובהק למדד ההעדפה pRight-pLeft (איור 2א, 2ב): limonene בהתאמה חיובית (לפי אחוז מסך נדיפים: p=0.0002 , R2=0.27; לפי ריכוז בעלווה: p=0.0003, R2=0.25), ו-a-pinene בהתאמה שלילית (לפי אחוז מסך נדיפים: p=0.0015 , R2=0.20; לפי ריכוז בעלווה:p=0.003 , R2=0.17).
הקשר בין מדדים תזונתיים לבין העדפה – נמצא כי להפרש בתכולת טָנינים (באחוזים ממשקל יבש) בין שיח ימין לשיח שמאל קשר ליניארי שלילי מובהק למדד ההעדפה pRight-pLeft (p<0.0001, R2=0.34) (איור 2ג). לא נמצא קשר בין מדד ההעדפה לבין ההפרשים בתכולות חלבון, מינרלים וסיבים ובנעכלות העלווה.
קשר משולב בין נדיפים בודדים וטָנינים לבין העדפה – השילובים בין טָנינים ל-limonene ובין טָנינים ל-a-pinene נמצאו בעלי קשר מובהק למדד ההעדפה pRight-pLeft (p<0.0001, R2=0.39 ו-p<0.0001, R2=0.41 בהתאמה). שני הגורמים המשתתפים תרמו באופן מובהק למודל הסטטיסטי בכל סידור שהוא בשני המודלים.
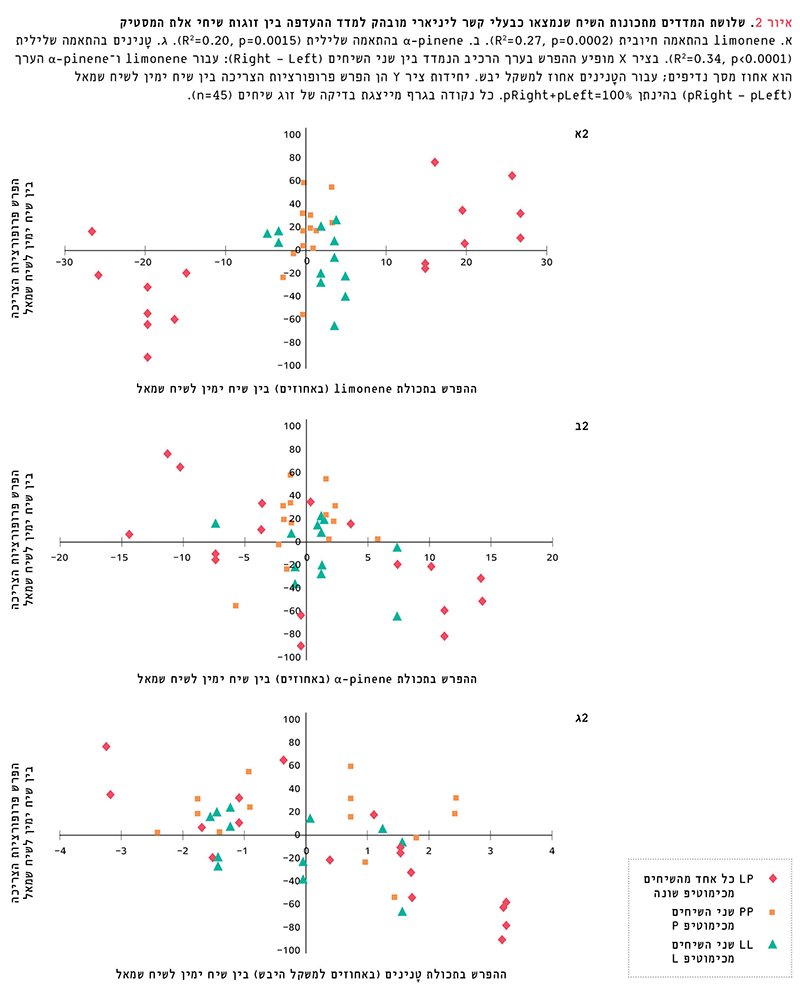
איור 2
שלושת המדדים מתכונות השיח שנמצאו כבעלי קשר ליניארי מובהק למדד ההעדפה בין זוגות שיחי אלת המסטיק
א. limonene בהתאמה חיובית (R2=0.27, p=0.0002). ב. α-pinene בהתאמה שלילית (R2=0.20, p=0.0015). ג. טָנינים בהתאמה שלילית (R2=0.34, p<0.0001). בציר X מופיע ההפרש בערך הרכיב הנמדד בין שני השיחים (Right – Left): עבור limonene ו-α-pinene הערך הוא אחוז מסך נדיפים; עבור הטָנינים אחוז למשקל יבש. יחידות ציר Y הן הפרש פרופורציות הצריכה בין שיח ימין לשיח שמאל (pRight – pLeft) בהינתן pRight+pLeft=100%. כל נקודה בגרף מייצגת בדיקה של זוג שיחים (n=45).
דיון ומסקנות
רעיית עדרי עיזים בחורש ים תיכוני מציעה פתרון יעיל למספר בעיות מהותיות בניהול שטחים טבעיים אלה [17, 32, 34]. בהקשר זה חשוב להזכיר שרבים משיחי החורש מופצים על-ידי ציפורים [29]. היות שכך, ייתכן כי לבית הגידול המצומצם יחדרו לאורך השנים זרעים ממקורות מגוונים שיגרמו להגדלת השונות הגנטית בין הפרטים באוכלוסיית המין. לכן, אנו סבורים כי בביצוע תצפיות על יחסי הגומלין בין אוכלי צמחים לעצים ולשיחים בחורש ראוי להיות ערניים גם לתבניות שיחור הבאות לביטוי ברזולוציה עדינה שבתוך רמת המין.
שאלתנו המרכזית במחקר זה הייתה: האם וכיצד תשפיע השונות הכימית הכוללת שנמצאה באוכלוסיית אלת המסטיק שבחלקת כבארה על התנהגות הרעייה של עדר עיזים שייכנס לרעות בחלקה? לשם כך ביצענו ניסוי העדפה בתנאים מבוקרים. תחילה בחנו אם לעיזים קיימת העדפה כלשהי בין שני טיפוסי הנדיפים הבולטים באוכלוסייה. התוצאות לימדו כי לעיזים אכן קיימת העדפה ברורה, בעוצמה בינונית, לשיחי אלת המסטיק מכימוטיפ L על פני שיחים מכימוטיפ P.
מגוון גורמים מתכונות השיח, נוסף על זהות כימוטיפ-הנדיפים, עשויים להיות מעורבים באירועי העדפה בין שיחי אלת המסטיק שכנים. ביניהם בולטים במיוחד הטָנינים, שתפקידם כרעלן מבוסס היטב [12, 19]. ואכן, במחקר נמצא קשר ליניארי שלילי מובהק בין תכולת טָנינים בעלווה לבין העדפה בין שיחים (איור 2ג). ניתוח שונות משותפת מצא כי זהות הכימוטיפ ותכולת הטָנינים הן צירוף התכונות הדומיננטי ביותר מבין כלל תכונות השיח המעורבות באירועי העדפה, ולשני הרכיבים תרומה מובהקת להסברת ההעדפה.
כאשר עוסקים בהשפעה אפשרית של פרופיל הנדיפים המרכיבים את השמן האתרי על העדפות של אוכלי צמחים, חובה עלינו לבחון גם נדיפים בודדים. שני רכיבים נדיפים נמצאו כבעלי קשר מובהק להעדפה בין שיחים: limonene בהתאמה חיובית ו-a-pinene בהתאמה שלילית (איור 2א, 2ב). התוצאות הללו תואמות למידע שקיים בספרות [16, 36], ומתלכדות עם הממצאים ברמת הכימוטיפ. השילוב של תכולת טָנינים (קשר שלילי) עם limonene (קשר חיובי) נמצא כגורם המסביר הפוטנציאלי הטוב ביותר להעדפה בין שיחי אלת המסטיק אצל עיזים.
הממצאים מלמדים כי עיזים במרעה חורש ים תיכוני ערות להבדלים בין פרטים שכנים של אלת המסטיק, והדבר מתבטא בהתנהגות הרעייה שלהן. ניתן להעריך כי העדפות בין צמחים שכנים מאותו המין קיימות גם אצל אוכלי צמחים נוספים הניזונים מאוכלוסיות מקומיות של מינים מעוצים אחרים בחורש. לכן, בביצוע תצפיות על הנעשה בחורש מומלצת ערנות גם למתרחש בתוך רמת המין.

המנגנון מאחורי הממצא
רכיבים משניים מספקים לצמחים הגנה מפני אכילה בשתי דרכי פעולה אפשריות השונות מאוד זו מזו: הגנה על-ידי פעולה פיזיולוגית ישירה של הרכיב כנגד אוכל הצמחים [26, 40, 42], והגנה עקיפה, שהרכיב מעורב בה באינטראקציה מערכתית ומורכבת יותר בין אוכל הצמחים, הצמח, ולעיתים גם אורגניזמים נוספים במערכת [27, 28, 35]. בין תפקודי ההגנה העקיפים המוכרים היטב בספרות: שחרור מקבץ נדיפים ייחודי על-ידי הצמח כקריאת מצוקה לגיוס אויבים טבעיים של אוכל הצמחים התוקף אותו [14], וחתימת טעם או ריח המעידה על איכות תזונתית ירודה [27]. במחקר זה הדגמנו מקרה שבו שני סוגי ההגנות, הישירה – הפיזיולוגית והעקיפה – המערכתית, פועלים בו-זמנית באלת המסטיק ומשפיעים על ההעדפות של העיזים.
טָנינים הם דוגמה טובה ומבוססת להגנה על-ידי פעולה פיזיולוגית ישירה [12, 19], אך שאלת מנגנון השפעת הנדיפים על ההעדפה שנצפתה, מסובכת הרבה יותר. נדיפים יכולים גם הם לגרום נזק פיזיולוגי ישיר לאוכלי צמחים [25, 40], אבל בו-בזמן משמשים נשאי מידע במערכת המקומית, ומספקים לאוכלי הצמחים מידע לגבי איכותו של הצמח כמקור מזון, על-ידי חישה באמצעות הטעם והריח [11, 13, 27, 28]. אם כן, מי משני מנגנונים אלה אחראי להעדפה שנצפתה בין שני הכימוטיפים L ו-P במחקר זה? נראה שההשפעה של הנדיפים במקרה שלפנינו אינה יכולה להיות מוסברת באפקט פיזיולוגי ישיר מהסיבות הבאות: א. סך ריכוז השמן האתרי באלות המסטיק שבחלקת כבארה היה נמוך (0.15% ממשקל יבש) [2]; ב. הטֶרפֵּנים הפחמימניים, שהם כ-95% מהשמן האתרי באלת המסטיק [3], נחשבים יעילים פחות לפגיעה ישירה באורגניזמים אחרים [16]; ג. בחישוב שהתבצע [בעקבות 30] נמצא כי הכמות הנמוכה של שמן אתרי שנמצאה באלת המסטיק אינה יכולה להגיע לריכוז שעלול לגרם נזק לפלורה החיידקית בכרס העז; ד. ליונקים מנגנונים יעילים לנטרול רעילות של טֶרפֵּנים [16], ומכאן שהשפעה ישירה על גוף העז עצמו גם היא אינה סבירה. מכל אלה, אנו מסיקים כי ההעדפה שנצפתה אינה יכולה להיות מוסברת על-ידי השפעה פיזיולוגית ישירה של הנדיפים על העז, אלא יש להסבירה במסגרת פעילותם של הנדיפים כנשאי מידע מתווכים במערכת התקשורת שבין האורגניזמים בבית הגידול.
חושי הטעם והריח משמשים אמצעי קלט יעיל לאוכלי הצמחים, ומספקים להם את היכולת לזהות, ללמוד ולזכור את האיכות התזונתית של צמחים שונים בתוך סביבה מורכבת ומשתנה תדיר [35]. באופן כללי, אוכלי צמחים יזהו את מכלול הטעם והריח של צמח בעל ערכים תזונתיים גבוהים כטוב, ואת הטעם והריח של צמח בעל איכות תזונתית נמוכה כרע [35]. אם כן, יש לשאול: האם העובדה שהטעם והריח של שיחי אלת המסטיק מכימוטיפ L מועדפים על העיזים יותר מאשר הטעם והריח של שיחים מכימוטיפ P, משמעותה שלראשון ערכים תזונתיים גבוהים יותר מאשר לאחרון? התוצאות שלנו מלמדות שהתשובה לשאלה זו שלילית. הכימוטיפיםL ו-P אינם שונים בפרמטרים התזונתיים שמדדנו: חלבון, סיבים, מינרלים ונעכלות. חשוב מכך, לא נמצא הבדל בין הכימוטיפים בתכולת הטָנינים, שהם ללא ספק הרעלן הבולט ביותר באלת המסטיק. עד כמה שאנחנו יכולים לקבוע, אין כל הבדל בערך התזונתי של הכימוטיפים L ו-P, ועדיין האחד מועדף על-ידי העיזים על פני האחר. מדוע?

ההסבר שאנו מציעים כאן מתבסס על העובדה שהמגע בין אוכלי צמחים לצמח מסוים הוא אף פעם לא אירוע בודד ומנותק מהקשר. במהלך שיחור המזון בשטחי המרעה נתקל אוכל הצמחים במיני צמחים רבים. נוסף על כך, המונו-טֶרפֵּנים הבסיסיים, כמו אלה הנמצאים באלת המסטיק, משותפים למספר רב של מיני צמחים. אף על פי שצמחים מייצרים עשרות ומאות נדיפים, הרי שחתימת הארומה של הצמח, העוזרת לאוכל הצמחים בבחירת המזון, מתבססת בדרך כלל על תת-קבוצה מצומצמת יחסית של נדיפים [21]. אם כן, ההסבר שאנו מציעים הוא שהעיזים באו כבר עם "דעה קדומה" לגבי a-pinene ושאר המונו-טֶרפֵּנים [3] שמאפיינים את כימוטיפ P וזאת בעקבות אינטראקציה עם מיני צמחים אחרים בשטחי המרעה בהווה או בעבר, או לחלופין בגלל המורשת הבין-דורית של העדר [7].
לסיכום: מרבית המחקרים שבחנו את תפקידם של נדיפים במערכת התקשורת שבין צמחים לאוכלי צמחים הדגישו את חשיבותם כנשאי מידע על הערך התזונתי של הצמח המשחרר אותם [28]. המסקנות שלנו מלמדות שגם במקרים שנדיפים אינם מכילים מידע אמיתי לגבי הערך התזונתי של הצמח, עדיין יכולה להיות להם השפעה על הנעשה בבית הגידול, וזאת כתוצאה מהפרשנות תלוית ההקשר שאוכל הצמחים מייחס להם. תמונת המצב המתקבלת היא של מערכת יחסים כאוטית ודינמית הצפויה לשמר מגוון גנטי. כדי להבין לעומק את ההשפעות ההדדיות בין צמחים לאוכלי צמחים על המתצפת לתת משקל גם לפרשנות שנותן אוכל הצמחים לנדיפים מהצומח. לכן, על האקולוג-החוקר או חובב הטבע לבחון ולהכיר את המערכת שלפניו באופן מקומי וברזולוציה מפורטת יותר, שמביאה בחשבון את ההיסטוריה של אוכלוסיית אוכל הצמחים בבית הגידול ואת מכלול מיני הצומח המרכיבים את מזונו באתר המקומי.
תודות
לצוות פארק הטבע רמת הנדיב, שהיה האכסניה למחקר – על שיתוף הפעולה הפורה, הרצון הטוב והסיוע. לאבי פרבולוצקי, שליווה את המחקר בעצות מועילות. ליאן לנדאו – על הייעוץ ועל הסיוע המעשי. לאלכסנדר ווינשטיין, הילרי פוט, תניה מסקי, בן רימון, אלונה שכטר ולבנה דבש – על הסיוע.
המאמר מוקדש לזכרו של מתניה שטרן, רועה צאן שהיה למורה, והדגים לתלמידיו חיים של חזון ואחריות.
- לתחום המחקר של רעיית בעלי חיים צמחוניים בחורש ים תיכוני חשיבות רבה בגיבוש מדיניות ניהול שטחי החורש.
בחינת דפוסי שיחור המזון של עיזים משפרת את הבנת השפעתה של הרעייה על ממשק החורש. - המחקר מצא שיש שונות בין פרטים בני אותו מין באוכלוסיית הצומח (במקרה זה – אלת המסטיק) שגורמת להבדלים בשיעור צריכתם על–ידי עיזים. ממצא זה מחדד את ההבנה לגבי העדפות פרטניות של חיות מרעה.
- המאמר מוסיף נדבך להבנת היבט אקולוגי המשפיע על רעיית עיזים בחורש, שלה חשיבות במניעת שרפות ובשמירה על מגוון בתי גידול לחיות הבר ולמיני הצומח.
המערכת
נספחים
נספח 1
להורדהמקורות
- כבביה ד. 1994. התנהגות רעיה והזנה של עיזים במרעה חורש ים-תיכוני (עבודת גמר לתואר מוסמך). רחובות: האוניברסיטה העברית בירושלים.
- נבון ש. 2014. מבט מקרוב על התנהגות הרעייה של עיזים בחורש ים תיכוני (עבודת גמר לתואר מוסמך). רחובות: האוניברסיטה העברית בירושלים.
- נבון ש, קיגל ח, דודאי נ ואונגר י"ד. 2016. שונות כימית באוכלוסייה מקומית של אלת המסטיק (Pistacia lentiscus L.). יער 16: 36–45.
- פרבולוצקי א. 1991. נקמת השעיר לעזאזל. טבע וארץ ל"ג: 4.
- שביב ע. אאוטאקולוגיה של אלת המסטיק (Pistacia lentiscus L.) (עבודה לקבלת תואר דוקטור). חיפה: הטכניון – מכון טכנולוגי לישראל.
- שמידע א. עושר המינים והאבולוציה של המינים החד-שנתיים בחורש הים-תיכוני. עלון רתם 18: 57–68.
- Arviv A, Muklada H, Kigel J, et al. 2016. Targeted grazing of milk thistle (Silybum marianum) and Syrian thistle (Notobasis syriaca) by goats: Preference following preconditioning, generational transfer, and toxicity. Applied Animal Behaviour Science 179: 53-59.
- Avrani S, Ben-Shlomo R, and Inbar M. 2012. Genetic structure of a galling aphid Slavum wertheimae and its host tree Pistacia atlantica across an Irano-Turanian distribution: From fragmentation to speciation? Tree Genetics & Genomes 8: 811-820.
- Baraza E, Hódar JA, and Zamora R. 2009. Consequences of plant–chemical diversity for domestic goat food preference in Mediterranean forests. Acta Oecologica 35: 117-127.
- Bashan D and Bar‐Massada A. 2017. Regeneration dynamics of woody vegetation in a Mediterranean landscape under different disturbance‐based management treatments.Applied Vegetation Science 20: 106-114.
- Bedoya-Pérez MA, Isler I, Banks PB, and McArthur C. 2014. Roles of the volatile terpene, 1, 8-cineole, in plant–herbivore interactions: A foraging odor cue as well as a toxin?. Oecologia 174: 827-837.
- Decandia M, Sitzia M, Cabiddu A, et al. 2000. The use of polyethylene glycol to reduce the anti-nutritional effects of tannins in goats fed woody species. Small Ruminant Research 38: 157-164.
- Delphia CM, Rohr JR, Stephenson AG, et al. 2009. Effects of genetic variation and inbreeding on volatile production in a field population of International Journal of Plant Sciences 170: 12-20.
- Dicke M and Baldwin IT. 2010. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends in Plant Science 15: 167-175.
- Hamrouni‐Aschi E, Larbi Khouja M, Boussaid M, et al. 2013. Essential oil composition of the Tunisian endemic cypress (Cupressus sempervirens var. numidica Trab.). Chemistry & Biodiversity 10: 989-1003..
- Estell RE, Fredrickson EL, Tellez MR, et al. 1998. Effects of volatile compounds on consumption of alfalfa pellets by sheep.Journal of Animal Science 76: 228-233.
- Evlagon D, Kommisarchik S, Glasser T, et al. 2010. How much browse is available for goats that graze Mediterranean woodlands?. Small Ruminant Research 94: 103-108.
- Glasser T, Landau S, Ungar ED, et al. 2008. A fecal near-infrared reflectance spectroscopy-aided methodology to determine goat dietary composition in a Mediterranean shrubland. Journal of Animal Science 86: 1345-1356.
- Glasser TA, Landau SY, Ungar ED, et al. 2012. Foraging selectivity of three goat breeds in a Mediterranean shrubland. Small Ruminant Research 102: 7-12.
- Goetsch AL, Gipson TA, Askar AR, and Puchala R. 2010. Feeding behavior of goats. Journal of Animal Science 88: 361-373.
- Goff SA and Klee HJ. 2006. Plant volatile compounds: Sensory cues for health and nutritional value? Science 311: 815-819.
- Korol L, Shklar G, and Schiller G. 2004. Tabor oak in Israel, genetic diversity within and between populations. Forest Genetics 11: 317-323.
- Landau S, Muklada H, Markovics A, and Azaizeh H. 2014. Traditional uses of Pistacia lentiscus in veterinary and human medicine. In: Yaniv Z and Dudai N (Eds). Medicinal and Aromatic Plants of the Middle-East. Netherlands: Springer.
- Llusia J and Penuelas J. 1998. Changes in terpene content and emission in potted Mediterranean woody plants under severe drought. Canadian Journal of Botany 76: 1366-1373.
- Macheboeuf D, Morgavi DP, Papon Y, et al. 2008. Dose-response effects of essential oils on in vitro fermentation activity of the rumen microbial population. Animal Feed Science Technology 145: 335-350.
- Makkar HPS. 2003. Effect and fate of tannins in ruminant animals, adaptation to tannins and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Ruminant Research 49: 241-256.
- Massei G, Cotterill JV, Coats JC, et al. 2007. Can Batesian mimicry help plants to deter herbivores? Pest Management Science 63: 559-563.
- Moore BD, Wallis IR, Palá-Paúl J, et al. 2004. Antiherbivore chemistry of Eucalyptus – Cues and deterrents for marsupial folivores. Journal of Chemical Ecology 30: 1743-1769.
- Nahum S, Inbar M, Ne’eman G, and Ben-Shlomo R. 2008. Phenotypic plasticity and gene diversity in Pistacia lentiscus along environmental gradients in Israel. Tree Genetics and Genomes 4: 777-785.
- Nagy JG and Tengerdy RP. 1968. Antibacterial action of essential oils of Artemisia as an ecological factor II. Applied Microbiology 16: 441-444.
- Ne’eman G. 1993. Variation in leaf phenology and habit in Quercus ithaburensis, a Mediterranean deciduous tree. Journal of Ecology 81: 627-634.
- Papachristou TG, Dziba LE, and Provenza FD. 2005. Foraging ecology of goats and sheep on wooded rangelands. Small Ruminant Research 59: 141-156.
- Peñuelas J and Llusià J. 2004. Plant VOC emissions: Making use of the unavoidable. Trends in Ecology and Evolution 19: 402-404.
- Perevolotsky A and Seligman NAG. 1998. Role of grazing in Mediterranean rangeland ecosystems. BioScience 48: 1007-1017.
- Provenza FD, Villalba JJ, Dziba LE, et al. 2003. Linking herbivore experience, varied diets and plant biochemistry diversity. Small Ruminant Reserarch 49: 257-274.
- Riddle RR, Taylor Jr CA, Kothmann MM, and Huston JE. 1996. Volatile oil contents of ashe and redberry juniper and its relationship to preference by Angora and Spanish goats. Journal of Range Management 49: 35-41.
- Rogosic J, Pfister JA, Provenza FD, and Grbesa D. 2006. Sheep and goat preference for and nutritional value of Mediterranean maquis shrubs. Small Ruminant Research 64: 169-179.
- Schiller G, Herr N, Shklar G, and Korol L. 2005. Short Communication: Diversity assessment of leaf phenology variation in Quercus ithaburensis Decne by RAPD. Israel Journal of Plant Sciences 53: 75-78.
- Stutz RS, Banks PB, Proschogo N, and McArthur C. 2016. Follow your nose: leaf odour as an important foraging cue for mammalian herbivores. Oecologia 182: 643-651.
- Utsumi SA, Cibils AF, Estell RE, et al. 2009. Seasonal change in one seed juniper intake by sheep and goats in relation to dietary protein and plant secondary metabolites. Small Ruminant Research 81: 152-162.
- Villalba JJ, Provenza FD, and Bryant JP. 2002. Consequences of the interaction between nutrient and plant secondary metabolites on herbivore selectivity: Benefits or detriments for plants? Oikos 97: 282-292.
- Villalba JJ, Provenza FD, and Olson KC. 2006. Terpenes and carbohydrate source influence rumen fermentation, digestibility, intake and preference in sheep. Journal of Animal Science 84: 2463-2473.


 מהדורה מודפסת
מהדורה מודפסת








