
חזית המחקר
שיטות פיזיולוגיות לקביעת עמידות של עצי אלון התבור וצאצאיהם ליובש
שמעון לביא הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושלים; המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקני
בני אבידן המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקני
סוהיל זיידן אגף הייעור, קרן קיימת לישראל
אביב אייזנבנד אגף הייעור, קרן קיימת לישראל
יאיר מני המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקני
יוסי יניב המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקני
איריס ביטון המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקני
רוני וולך הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושלים
מנחם מושליון הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושלים
גיורא בן-ארי המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקני ([email protected])

שלבים בנביטת בלוט של אלון התבור | צילום: היא-לי בונפיל
היא-לי בונפיל
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקנישמעון לביא
הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושלים; המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניבני אבידן
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניסוהיל זיידן
אגף הייעור, קרן קיימת לישראלאביב אייזנבנד
אגף הייעור, קרן קיימת לישראליאיר מני
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקנייוסי יניב
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניאיריס ביטון
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקנירוני וולך
הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושליםמנחם מושליון
הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושליםגיורא בן-ארי
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניהיא-לי בונפיל
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקנישמעון לביא
הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושלים; המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניבני אבידן
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניסוהיל זיידן
אגף הייעור, קרן קיימת לישראלאביב אייזנבנד
אגף הייעור, קרן קיימת לישראליאיר מני
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקנייוסי יניב
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניאיריס ביטון
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקנירוני וולך
הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושליםמנחם מושליון
הפקולטה לחקלאות רחובות, האוניברסיטה העברית בירושליםגיורא בן-ארי
המכון למדעי הצמח, מנהל המחקר החקלאי – מרכז וולקניבעקבות תחזיות אקלים המנבאות שינויים בתדירות המשקעים ופריסתם והתחממות, ובעקבות דיווחים על אודות התייבשות עצי יער בארץ ובעולם, גובר הצורך בהבנת מנגנוני הישרדות של עצים ביובש ובמציאת טיפוסים עמידים ליובש. כיום מרבים את אלוני התבור במשתלות קק"ל על-ידי איסוף בלוטים מעצים שונים באזורים שונים. בחירת עצי האֵם המספקים בלוטים אינה מבוססת על מאפיינים תפקודיים של העצים עצמם, מלבד מספר צאצאים גבוה וגישה נוחה. במחקר זה בחנו עצי אֵם בשטח, וכן את צאצאיהם בניסוי יובש במערכת גרוימטרית סגורה ('ליזימטרים'), ואפיינו את הקשר הגנטי ביניהם לקביעת אופן ההפריה. השערת המחקר שלנו היא כי בחירת עצי אם מושכלת יותר ומבוססת על פרמטרים פיזיולוגיים-גנטיים תביא לריבוי אלוני התבור מותאמים יותר לסביבתם בארץ, דבר שישפר את שרידותם ביערות קק"ל.
ממצאי הניסוי שערכנו מראים כי צאצאים מעצי אֵם בתנאי מחיה גבוליים הראו התנהגות איזוהידרית שמרנית (סגירת פיוניות בתגובה לעקת מים בקרקע). נראה שהתנהגות זו מעניקה יתרון ביובש קיצוני, מאחר שהיא מתאפיינת בשמירה על מוליכוּת פיוניות נמוכה ובהפסד מים מועט בדיוּת. עם זאת, קצב הגדילה של צמחים אלה היה אִטי ביותר, ועל כן יש להם סיכון אקולוגי בתחרות על אור ועל משאבים. צאצאים מעצי אֵם בתנאי מחיה טובים הראו התנהגות אנאיזוהידרית (עיכוב בתגובת הפיוניות לירידה בפוטנציאל המים בקרקע), ועל כן צמחו בקצב המהיר ביותר, אך תגובתם ליובש הייתה אִטית, ולכן התייבשותם הייתה דרמטית וחלק מהם לא שרדו בעקבות יובש.
צאצאי העצים מ'גבעת חציר', 'פרדס חנה' ו'כפר חנניה' הראו התנהגות שמרנית קיצונית, וצאצאי העץ ב'כחל' הראו התנהגות דומה אך מתונה יותר. לכן, אנו ממליצים על עצים אלה כעצי אֵם לריבוי לשם נטיעה ביערות קק"ל. צאצאי העצים 'טייבה' ו'כפר יונה' הראו צמיחה מהירה, אך ניחנו בהתנהגות מסתכנת, ולכן אנו ממליצים על שימוש בעצים אלה באזורים גאוגרפיים רוויי משקעים.
על קצה המזלג
- התמודדות עם התייבשות עצים היא מאתגרי היערנות המרכזיים לנוכח שינוי האקלים והדיווחים על תמותת עצים ברחבי העולם. יש צורך מידי בגיבוש תכנית להשבחת עצים עמידים ליובש לצורך נטיעות.
- הבנת הבסיס הגנטי של אוכלוסיות עצים הכרחית להכנת תכנית שכזו, ובפרט כשמדובר בעצי יער שהידע הגנטי על אודותיהם מוגבל. התכנית יכולה לסייע בבחירת עצים שישמשו כמקור לזרעים.
- אלון התבור הוא מין מקומי וחשוב בייעור הארץ. מסתבר כי בין אלוני התבור הגדלים בבתי גידול שונים, ואף בין כאלה הגדלים באותו בית גידול, ישנם הבדלים ניכרים בהתנהגות הפיזיולוגית בתנאי יובש.
- המאמר מקדם ההבנה על אודות אופי הורשת תכונת העמידות ליובש באלוני התבור – תכונה חשובה לתכנון נטיעת יערות העתיד.
המערכת
הלכה למעשה
מבוא
מיני הסוג 'אלון', Quercus, הם 'מין מפתח' בבתי הגידול השונים שלהם ברחבי העולם. ה'אלון' נפוץ בטווח רחב של תנאי אקלים, החל באזורים ים תיכוניים צחיחים למחצה ועד יערות הגשם התת-טרופיים [25]. צפיפותם הולכת וקטנה ברחבי העולם בעקבות השפעות אקלים קיצוניות (עקות, צינה ויובש) [11, 26].
אלון התבור (Quercus ithaburensis [Decne]) ממשפחת האלוניים (Fagaceae), הוא אחד משלושת מיני האלון הגדלים בישראל (למעט אזור החרמון). אלון התבור מפתח נוף גדול ורחב על גזע מרכזי, ומגיע לגובה של 15 מטר [6, 8]. כמין אירנו-טורני שעבר התאמה לאקלים הים תיכוני [16, 20, 24], גדל האלון בעיקר כיער פארק בשילוב צמכחייה רב-שנתית כמין יחיד או כמלווה מינים אחרים [6, 8]. הסוג 'אלון' בכלל, והמין אלון התבור בפרט, מאופיין בשונות צורנית והתפתחותית גדולה המתבטאת בצורת הספלול והבלוט, בצורת העלים ובמועדי הלבלוב והפריחה [26, 31]. שונות זו מיוחסת לשינויי האקלים במזרח הים תיכון [21] וכן לשונות הגנטית במיני האלון [1, 14]. בחינה של אוכלוסיית אלון התבור בארץ בעזרת סמני די-אן-איי חשפה שהיא מתחלקת לשלוש תת-אוכלוסיות בהתאם למיקומן הגאוגרפי – אוכלוסיית הגולן, אוכלוסיית הגליל התחתון ואוכלוסיית מישור החוף [24]. מקובל לחשוב כי קיים אי-התאם עצמי באלונים[15], ועל כן חצי מהדי-אן-איי של הצאצאים – מקורו זר .
ריבוי אלון התבור במשתלות קק"ל נעשה על-ידי הנבטת פֵּרותיו. ריבוי צמחי נוסה פעמים רבות ללא הצלחה מסחרית [1] ולכן אינו משמש ליצירת עצים חדשים. לשם נטיעות חדשות של עצי אלון התבור ביערות קק"ל נאספים בכל שנה בלוטים של אלון התבור מעצים שונים באוכלוסיות שונות. הבלוטים נאספים מעצים שהגישה אליהם נוחה, אך לא בהכרח מעצים עמידים ליובש.
במחקריו של הר [4, 5] נמצא כי משק המים הוא גורם מגביל בהתפתחות ובתפוצה של אלוני התבור בישראל. במחקר שערך קופר [7] נמצא כי באביב התקבלו במין זה הערכים הפיזיולוגיים הגבוהים ביותר (בשיא עוצמת הקרינה), ובסוף הקיץ נצפה ערך קיצון של פוטנציאל המים בגזע (Ψw) שהגיע ל-MPa (-3.5) (ערכים דומים נצפו גם במחקרו של הר). נוסף על כך, Ψw נמוך מ-(2.8-) MPa הביא לסגירת הפיוניות ולהטמעה שלילית, ונטען כי אלון התבור מתנהל בצורה איזוהידרית (isohydric) להתמודדות עם יובש (בעלי ניהול מים "שמרני"). לעומתו, לאלון המצוי נטייה לאנאיזוהידריות, ומוליכות הפיוניות נשמרת גם בפוטנציאל מים [9] של (-4) MPa. בדרך כלל לכל מין יש התנהגות טיפוסית כתגובה ליובש, אך יכולים להיות הבדלים גם בין טיפוסים מאותו המין, כפי שנצפה בצפצפות [10] ובאלונים [12]. ישנם חילוקי דעות בשאלה איזו התנהגות היא בעלת יתרון גדול יותר בעמידות ליובש אצל צמחים-רב שנתיים [19, 22, 23], כיוון שצמחים איזוהידריים נמנעים מסיכון הידראולי אך מסתכנים בהרעבת פחמן [19].
בעקבות תחזיות האקלים המנבאות צחיחות והתחממות [27] ולנוכח אופן ריבוי אלון התבור במשתלות קק"ל, מטרת מחקר זה היא מציאת טיפוסי אלון התבור המעמידים צאצאים בעלי יתרון ביובש.
שיטות וחומרים
כדי לאתר טיפוסים של אלון התבור שמעמידים צאצאים בעלי יתרון ליובש, נערכו שלושה אפיונים המשלימים זה את זה:
עצי אֵם
נבחרו 13 עצים בוגרים של אלון התבור על פי אופן החלוקה הבא: עצים עתיקים: טייבה (T), פרדס חנה (P), אלונים (A), כפר חנניה (KH), הגושרים (G) וחורשת טל (HT) [2]; עצים המשמשים את קק"ל לאיסוף בלוטים: כפר יונה (KY), כחל (K); חמישה עצים מאוכלוסייה טבעית בגבעת חציר הנמצאת בתנאי יובש [4, 5] וקרובים לעצים יבשים (GH 1,2,3,6,8).
פוטנציאל המים בגזע (Ψw) נמדד בעזרת תא לחץ (3000 ARIMAD של חברת MRC בישראל). מדידות פוטנציאל המים נעשו בשני מועדים: א. טרום-שחר (predawn, Ψpd), משקף את מצב המים ההתחלתי של אותו היום בקרקע; ב. צהריים (midday, Ψm), משקף את פוטנציאל המים בשיא הפעילות הפיזיולוגית על פי מדידות מקדימות שנעשו על אלון התבור (הנתונים לא מופיעים). נעשו 4–10 חזרות לעץ.
במדידות עצי האֵם יש שונות גדולה מאוד בתנאי בית הגידול, בגילים ובמועדי המדידות, שונות שעשויה להוביל למסקנות לא מדויקות. לא נעשו מדידות שטח על עצים A ו-P בעקבות בעיות טכניות בשטח. פוטנציאל המים של עצי גבעת חציר חושב כממוצע אחד בהשוואה לשאר עצי האֵם.
ניסוי יובש
צמחי הניסוי
בלוטים נאספו מכל עץ אֵם שהשתתף בניסוי במהלך החודשים אוקטובר ונובמבר 2013. כל מקבץ בלוטים מאותו עץ אֵם נזרע בעציץ נפרד עם ורמיקוליט גודל 3 (אגריקל תעשיות הבונים, ישראל). לאחר חודשיים נבטו רוב הבלוטים (אמצע ינואר 2014), ואז הועברו לבית זכוכית כדי להאיץ את גדילתם. כיוון שמקור הבלוטים בעצים שונים, התקבלו הבדלים באחוזי הנביטה, ולכן מספר החזרות לכל עץ אֵם היה שונה: אוכלוסיית גבעת חציר (GH): שמונה צאצאים (מ-4 עצים שונים); אלונים (A): ללא צאצאים; הגושרים (G): תשעה צאצאים; חורשת טל (HT): שנים-עשר צאצאים; כחל (K): שמונה-עשר צאצאים; כפר חנניה (KH): שמונה צאצאים; כפר יונה (KY): שלושה-עשר צאצאים; פרדס חנה (P) עשרה צאצאים; טייבה (T) ארבעה-עשר צאצאים.
מהלך הניסוי
הניסוי נערך בחודשים יוני–יולי 2014 בחממה מבוקרת חלקית בפקולטה לחקלאות ברחובות מהאוניברסיטה העברית בירושלים (בשיתוף המעבדה של פרופ' מנחם מושליון ופרופ' רוני וולך). צינון החממה נעשה בעזרת מזרן לח. הטמפרטורה והלחות היחסית נעו בין 400–30 צלזיוס ו-55%–35 בהתאמה. מערכת המדידה כוללת 96 יחידות שקילה לפי הזמנה (לרית מדידות בע"מ, ישראל) השוקלות באופן רציף (כל 10 שניות) כל שתיל במערכת ליזימטרים. עציצי 3.9 ליטר שקולים מולאו ב-1,700 גרם מצע גידול בנטל 11 (טוף מרום גולן, ישראל). כשלושה חודשים לפני תחילת הניסוי הועתקו כל השתילים לעציץ הליזימטר כדי שיספיקו לפתח מערכת שורשים בכל נפח העציץ. כשבועיים לפני תחילת הניסוי הועברו העציצים לחממת הניסוי להתאקלמות. הניסוי החל כשהשתילים היו בני חצי שנה, ונערך בין התאריכים 3.6.2014–29.7.2014. בתחילת הניסוי צולם כל שתיל על רקע שחור, והונח על המשקל המיועד לו. הניסוי בוצע בו-זמנית על 92 שתילי אלון התבור שהונחו בארבעה בלוקים באקראי על גבי שולחנות הניסוי. הניסוי כלל שלושה שלבים: א. השקיה מיטבית; ב. יובש; ג. התאוששות.
חישוב של קוטר ושטח עלווה
קוטר השתילים נמדד בתחילת הניסוי ובסופו על-ידי קליבר דיגיטלי (קוטר). צילום העציצים נעשה בעזרת מצלמת ריפלקס (Canon EOS 650D, יפן) וחצובה ממרחק קבוע, ונערך בשלושה מועדים: היום הראשון לניסוי, היום האחרון ליובש, היום האחרון לניסוי. העציץ הונח על רקע שחור וליד סרגל מדידה. שטח העלווה חושב מצילומי העציצים ביום הראשון של הניסוי בעזרת תוכנת פוטושופ (Photoshop CS6). שטח העלווה נקבע בהתאם ליחס הפיקסלים בין העציץ לבין גודל ידוע. כדי לקבוע את מִתאם שטח העלווה בעזרת צילום לעומת שטח העלווה האמִתי, צולמו 30 צמחים שלא השתתפו בניסוי, ועליהם הוסרו לצורך מדידת שטח העלווה במכשיר סורק עלים LI-3100 Area Meter (LI-COR, נברסקה, ארה"ב). התוצאות הושוו לתוצאות מהצילום, וחושבה משוואת הרגרסיה שעל פיה תוקן שטח העלווה של כל העציצים שהשתתפו בניסוי (P>2.55X10-19, R2=0.95)
חישוב פרמטרים פיזיולוגיים
ניתוח נתוני משקל העציץ נערך באמצעות אלגוריתם שפיתחו פרופ' וולך ופרופ' מושליון [30]. האלגוריתם שוכלל והוסב לקוד Matlab (MathWorks, ארה"ב) על-ידי הלפרין [3]. הפרמטרים שנבחנו: תכולת המים המחושבת בקרקע (VWC), קצב הדיות מנורמל לשטח העלווה לכלל הצמח ( להלן – דיות, E) ומוליכות הפיוניות מנורמלת לשטח העלווה לכלל הצמח (להלן מוליכות, Gs). חישובי הפרמטרים תוקנו לשטח העלווה באמצעות משוואת הרגרסיה (ראו "חישוב קוטר ושטח עלווה").
שונות גנטית
במחקר נבחנה השונות הגנטית באמצעות סמני SSR (Simple Sequence Repeat) שהראו רמות גבוהות של פולימורפיזם [13, 28], ונבדק אם קיימת הפריה עצמית אצל אלון התבור.
הפקת הדי-אן-איי מעלי אלון התבור התבססה על הפקה מעלי גפן [17]. האפיון הגנוטיפי נעשה על-ידי סמני SSR פלואורסצנטיים שאופיינו במיני אלון אחרים [13, 28]. תוצרי רֵאקציות ה-PCR נשלחו למרכז הלאומי לטכנולוגיות גנומיות (הפקולטה למדעי הטבע, האוניברסיטה העברית בירושלים). בחינת גודל התוצר נעשתה באמצעות 3730xl DNA Analyzer (של חברת Applied Biosystems), וגודל התוצר נקבע באמצעות התוכנה Sequence ScannerTM v1.0. תוצאות האפיון הגנוטיפי נותחו באמצעות התוכנות: Microsat, PHYLYP ו-TreeView. על בסיס המרחקים הגנטיים בין הפרטים השונים נבנו עץ פילוגנטי לעצי האֵם ועץ לעצי אֵם ולצאצאים.
תוצאות
מדידות עצי האֵם
מדידות פוטנציאל המים שנערכו לאלוני התבור באזורים השונים בחודשים מאי ואוקטובר 2014, שימשו להערכת מצב המים בכל עץ, שכן ישנם עצים היושבים על מים רבים וישנם עצים המקבלים את מי הגשמים בלבד: העץ T (העתיק ביותר) נמצא בתוך מטע גויאבות ועל כן מושקה ברוב ימות השנה, כמו גם העץ KY שנמצא ליד פרדס מושקה. לעצים בצפון הארץ, HT ו-G, אין, כנראה, בעיות מים כיוון שהם קרובים מאוד לנחלי האזור. העצים P ו-KH אינם במצב טוב במיוחד: העץ P בעל סדק גדול במרכז הגזע ו-KH חציו יבש לגמרי. למעשה, לעצים P, KH, K ו-GH אין מקור מים קרוב. יש מתאם ברור בין מצב הבלוטים לפוטנציאל המים בעץ: כאשר העץ במצב "טוב" הוא יניב כמות בלוטים גדולה ובעלי אחוז נביטה גבוה (ראו נספח 1).
בניתוח שונות דו-גורמי לבחינת השפעת העץ וחודש המדידה על מדדי פוטנציאל המים, מתקבלת אינטראקציה מובהקת בין שני הגורמים (P<2.7e-14) (טבלה 1). נמצא שעם התקדמות העונה יורדים Ψpd ו-Ψm באופן מובהק, למעט בעץ T שאין בו הבדל ב-Ψpd בין החודשים מאי לאוקטובר, והעץ KH שאין אצלו הבדל ב-Ψm בגזע בין החודשים. כאשר בוחנים את ההפרש בין פוטנציאל המים לפנות בוקר לעומת הצהריים נראה כי בעצים GH, Kו-KH מתקבלים ערכים נמוכים וללא הבדל בין סוף האביב (מאי) לסוף הקיץ (אוקטובר).
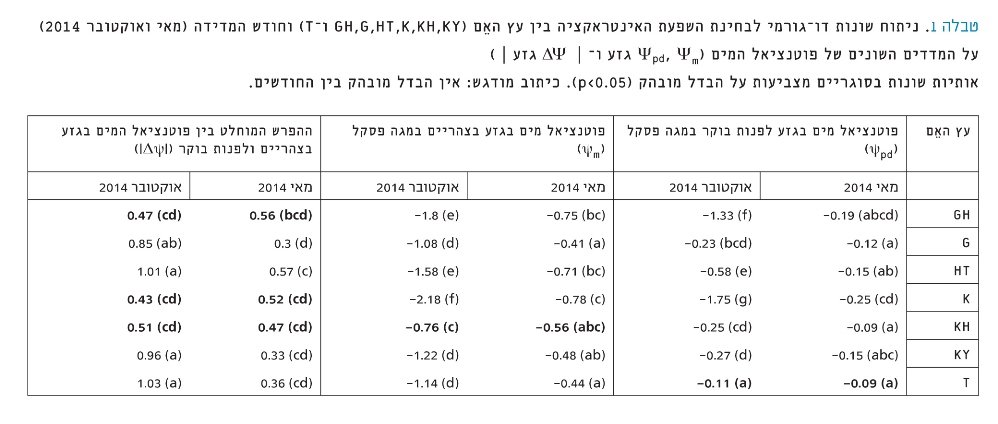
טבלה 1
ניתוח שונות דו-גורמי לבחינת השפעת האינטראקציה בין עץ האֵם (GH ,G ,HT ,K ,KH ,KY ו-T) וחודש המדידה (מאי ואוקטובר 2014) על המדדים השונים של פוטנציאל המים (Ψpd ,Ψm גזע ו–│ ΔΨ גזע │)
אותיות שונות בסוגריים מצביעות על הבדל מובהק (p<0.05). כיתוב מודגש: אין הבדל מובהק בין החודשים.
ניסוי יובש בצאצאי עצי האם
כל צאצאי עצי האֵם גודלו בחממה באותו זמן, בארבעה בלוקים באופן אקראי.
השוואה בין תכונות נמדדות בתנאים מיטביים
השפעת עץ האֵם על מדדי הצמיחה ועל מדדים פיזיולוגיים נבחנה בתנאי השקיה מיטביים (איור 1). צאצאי T הראו ערכים גבוהים באופן מובהק בכל המדדים. צאצאי KH, P ו-GH הציגו מדדי צמיחה נמוכים ביותר, וכן מוליכות פיוניות נמוכה וקצב דיות נמוך. לשלוש קבוצות הצאצאים של K, HT ו-G ערכים דומים בכל המדדים. ניתן לחלק באופן כוללני את צמחי הניסוי לשלוש קבוצות לפי מדדי הצמיחה שלהם ביום הראשון לניסוי: KY ו-T גדולי ממדים; HT, G ו-K בממדי ביניים; GH, KH ו-P בממדים הקטנים ביותר.
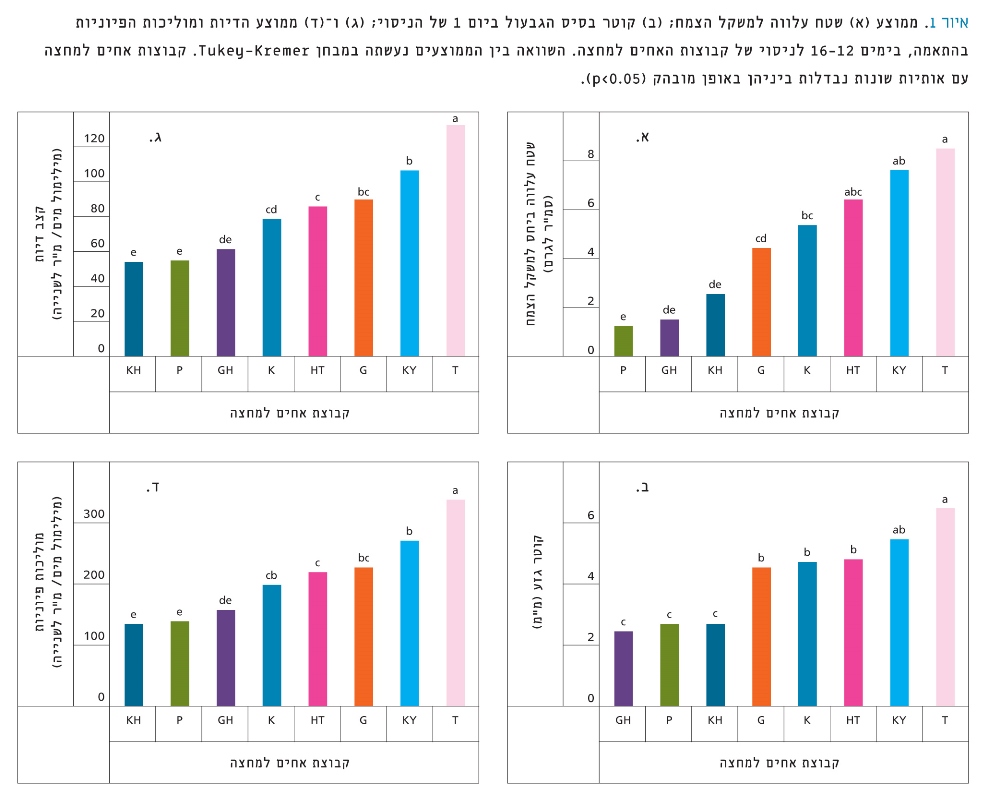
איור 1
ממוצע (א) שטח עלווה למשקל הצמח; (ב) קוטר בסיס הגבעול ביום 1 של הניסוי; (ג) ו-(ד) ממוצע הדיות ומוליכות הפיוניות בהתאמה, בימים 12–16 לניסוי של קבוצות האחים למחצה. השוואה בין הממוצעים נעשתה במבחן Tukey-Kremer. קבוצות אחים למחצה עם אותיות שונות נבדלות ביניהן באופן מובהק (p<0.05).
רגרסיית יובש והתאוששות – לכל קבוצת אחים למחצה חושבה רגרסיית היובש (ערכי דיות כתלות באחוז רטיבות הקרקע) ורגרסיית ההתאוששות (ערכי דיות לעומת ימים בהתאוששות) ונבחנו ההבדלים בין מקדמי הרגרסיה (טבלה 2). ברגרסיית היובש מתקבלים שיפועים שליליים עם הבדלים מובהקים ושונים מאפס. ככל שמתקדמים ביובש יש פחות דיות, ולצאצאי T השיפוע השלילי ביותר. שיפוע רגרסיית ההתאוששות של צאצאי KH, P ו-GH אינו שונה מאפס, ולכן ניתן לומר כי אין שינוי בדיות במהלך ההתאוששות. בין שאר קבוצות האחים למחצה (KY, K, HT ו-G) אין הבדל מובהק בין השיפועים, ולכן ניתן לומר כי כולם מתאוששים באופן דומה, אך מגיעים לדיות המיטבי שלהם באופן שונה. בהנחה כי רגרסיית ההתאוששות ממשיכה במגמה לינֵארית, חושבו הימים להתאוששות מלאה על פי משוואת הרגרסיה לכל קבוצת אחים למחצה, כאשר התאוששות מלאה חושבה כהגעה חזרה לערכי הדיות המרביים לפני היובש (טבלה 2). למרות האי-דיוק שבחישוב מכיוון שנעשה בצורה מעריכית, ניתן לראות שצאצאי G הגיעו להתאוששות מלאה ב-20.4 ימים, ומכאן להעריך כי אחריהם יתאוששו K ב-28.3 ימים, ו-KY ו-T יתאוששו בצורה מלאה רק לאחר 59.7 ו- 67.8 ימים, בהתאמה.
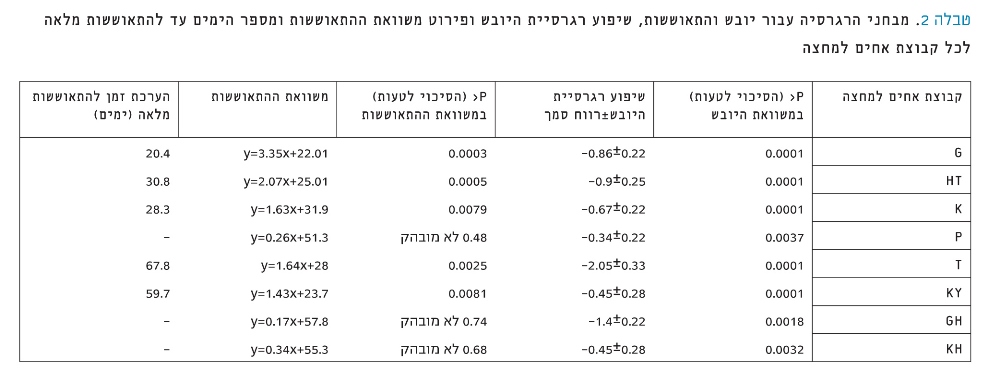
טבלה 2
מבחני הרגרסיה עבור יובש והתאוששות, שיפוע רגרסיית היובש ופירוט משוואת ההתאוששות ומספר הימים עד להתאוששות מלאה לכל קבוצת אחים למחצה
השוואה פרטנית בין צמחים בעלי שטח עלווה שונה – מוליכות הפיוניות אצל צמחים בעלי שטח עלווה שונה הצביעה על תלות בין המוליכות לשטח העלווה (איור 2). להדגמה, נבחרו שני צמחים מייצגים באופן הבא: צמח בעל שטח עלווה קטן, גדילה אִטית מאוד אך שלא נפגע ביובש (8-1-GH); לעומת צמח בעל שטח עלווה גדול מאוד, שנפגע ביובש והתאוששותו אִטית (10-KY). בשיא היובש איבד הצמח 10-KY 100% מהעלים, ואילו 8-1-GH לא איבד עלים כלל (הנתונים לא מופיעים). בשני הצמחים, ובכל שלבי הניסוי, שיא המוליכות התרחש בשעות הבוקר. 8-1-GH, בהשקיה מיטבית, מאופיין בשיא מוליכות נמוך באופן מובהק מ-10-KY, ובשלב היובש בעל מוליכות גבוהה באופן מובהק אחרי שעות הבוקר. בסוף היובש הצמח 10-KY בעל מוליכות פיוניות אפסית (למעט "חורים" קטנים במהלך היום), ובהתאוששות נראה שיש שיא בבוקר אך בשאר היום המוליכות אינה שונה באופן מובהק משלב היובש. אם כן, נראה שהצמח 10-KY נפגע מאוד במהלך היובש. הצמח 8-1-GH הוא בעל מוליכות פיוניות נמוכה אך קבועה לאורך כל שעות היום ולאורך כל שלבי הניסוי (למעט בין השעות 13:00–16:00 שהמוליכות ירדה בהן באופן מובהק לעומת ההשקיה המיטבית). נראה שאף על פי ש-8-1-GH לא איבד את עליו, ישנה ירידה במוליכות בזמן היובש וההתאוששות. כנראה, מדובר בחלק ממנגנון איזוהידרי לשמירה על תכולת מים קבועה בעלה על-ידי הורדת מוליכות הפיוניות בתנאי יובש.
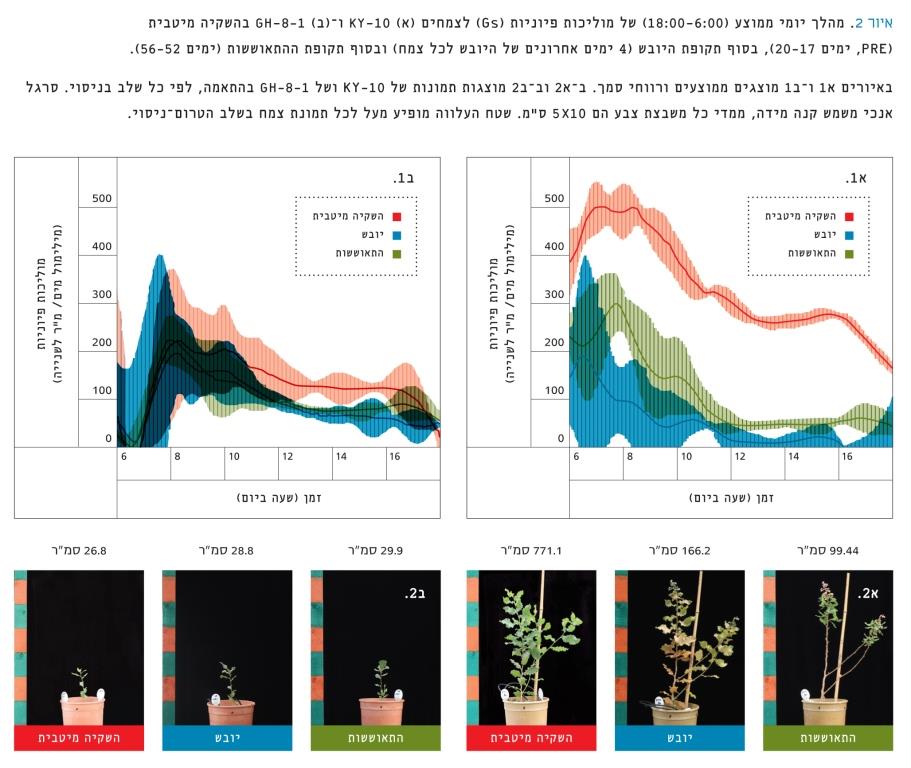
איור 2
מהלך יומי ממוצע (18:00-6:00) של מוליכות פיוניות (Gs) לצמחים (א) KY-10 ו-(ב) GH-8-1 בהשקיה מיטבית (PRE, ימים 17–20), בסוף תקופת היובש (4 ימים אחרונים של היובש לכל צמח) ובסוף תקופת ההתאוששות (ימים 52–56).
באיורים א1 ו-ב1 מוצגים ממוצעים ורווחי סמך. ב-א 2 וב-ב 2 מוצגות תמונות של KY-10 ושל GH-8-1 בהתאמה, לפי כל שלב בניסוי. סרגל אנכי משמש קנה מידה, ממדי כל משבצת צבע הם 5X10 ס"מ. שטח העלווה מופיע מעל לכל תמונת צמח בשלב הטרום-ניסוי.
שונות גנטית
על פי 11 סמני ה-SSR (נספח 2א) חושבו הקשרים הפילוגנטיים של עצי האֵם (נספח 2ב). ערכי ה-bootstrap (הבוחנים את המהימנות הסטטיסטית של עץ פילוגנטי) נמוכים עקב מספר קטן של סמנים. לפיכך, יש להתייחס לתוצאות אנליזה זו בעירבון מוגבל, מכיוון שייתכן שאינה משקפת נכונה את הקִרבה הגנטית בין העצים הנבחנים. קיימים צמדי עצים קרובים גיאוגרפית שקרובים גם מבחינה גנטית: זוגות העצים GH-1+GH-3, GH-8+GH-6, ו-KY+T. כאמור, בניתוח התוצאות התייחסנו לצאצאי העצים בגבעת חציר כאל אוכלוסייה למרות ההבדלים הגנטיים ביניהם.
לכל עץ אֵם נבחרו עד 5 צאצאים באופן אקראי, שעברו אפיון גנוטיפי בעזרת ארבעת הסמנים הפולימורפיים ביותר. נבחנו הקשרים הפילוגנטיים של עצי האֵם עם צאצאיהם (נספח 2ג). בסך הכול מדובר ב-11 עצי אֵם וב-44 צאצאים. צאצאי P ו-K קרובים להוריהם, לעומת KH, G ו-KY שנראה בהם מרחק גנטי גדול בין ההורים לצאצאיהם ובין הצאצאים השונים.
לכל פרט שנבדק התקבלו 8 אללים מ-4 סמנים שונים. במקרה של הפריה זרה, נצפה לפחות לאלל אחד בכל אתר שיהיה זהה לעץ האֵם (האלל שהורש מהאֵם). במקרה של הפריה עצמית נצפה לכלל האללים שיגיעו מעץ האֵם (ישנו סיכוי למוטציה במעבר לדור הצאצאים, אך באחוזים נמוכים). כל הצאצאים הכילו לפחות אלל אחד שככל הנראה לא הגיע מעץ האֵם (לא מוצג), ולכן ניתן להניח כי אצל אלוני התבור הנבחרים לא מתקיימת הפריה עצמית.
את תוצאות גודל הסמנים בזוגות בסיסים לעצי האם ולצאצאים ניתן לראות בנספח 3א ו-3ב, בהתאמה.
דיון ומסקנות
העצים שנבחרו לשמש עצי אֵם נבחרו מתוך מחשבה כי לעצים מבוגרים מאוד יש, כנראה, יתרון בעמידות לעקות אביוטיות וביוטיות, ועל כן יוכלו לשמש מקור זרעים מגוון לבחירת צאצאים עמידים ליובש.
בבדיקות פוטנציאל המים בגזע מתקבלים הבדלים בין העצים השונים: מבחינת Ψpd, נראה כי על פי הצפוי, ישנה ירידה לאורך העונה – במאי 2014 התקבל Ψpd גבוה באופן מובהק מזה שבאוקטובר 2014 (טבלה 1), למעט בעץ T, כיוון שהוא ממוקם ליד מים זמינים. העץ הוא המבוגר ביותר (נספח 1). לכן, ייתכן שנחשף לתנאים מיטביים לאורך תקופה ארוכה. מכאן ניתן להניח כי התנהגותו האנאיזוהידרית שימשה לו יתרון בסביבה רוויית מים זו. ב- Ψmנצפית בגזע מגמה דומה של העצים, אך הפעם אין הבדל מובהק בין החודשים בעץ KH. נראה כי לעץ KH התנהגות איזוהידרית: בעונה היבשה (אוקטובר 2014) מדד ה-Ψpd נמוך באופן מובהק מזה שנמדד במאי 2014, ושאר מדדי Ψ נשארים ללא שינוי (Ψm+Ψ∆ בגזע). תכולת המים בעלים בעץ KH נשארת גבוהה, כנראה, על-ידי הקטנת חילופי הגזים (מוליכות פיוניות נמוכה באוקטובר 2014, הנתונים לא מופיעים), אולם לא נערכו בדיקות פיזיולוגיות מעמיקות לאישוש הדבר.
במחקר של אשד [1] נמצא כי אין השפעה לגודל הבלוט על קצב הנביטה, אולם היה יתרון בולט לגודל הבלוט בצימוח הנבט. נתוני משקל הבלוטים חסרים, אך ניתן לראות כי המשקל ההתחלתי תואם לתיאור עצי האֵם: צאצאי T צמחו בקצב המהיר ביותר, ובתחילת הניסוי היו בעלי מדדי הצמיחה הגבוהים ביותר (איור 1). לעומתם, לצאצאי העצים ללא מקור מים זמין, P, KH ו-GH, היו מדדי הצמיחה הנמוכים ביותר. תוצאות דומות התקבלו במחקר שבחן את השפעת בית הגידול על גודל הבלוטים והשתילים ב-14 מיני אלון מצפון אמריקה [18]. נמצא כי צאצאי אלונים הגדלים בסביבה יבשה מאופיינים בשתילים הקטנים ביותר. צאצאי אלונים הגדלים בסביבה לחה הראו משקל נצר גדול יותר ביחס למשקל השורש, וגובה רב יותר ביחס למשקל השתיל לעומת האלונים מהסביבה היבשה.
חלק חשוב בניסוי זה היה בחינת אופן התאוששות הצמחים מניסוי היובש. התאוששות הוגדרה כחזרה לערכי הדיות לפני ניסוי היובש, כיוון שבניסוי זה לא קיימת קבוצת ביקורת. כדי להבין את אופן ההתאוששות של כל קבוצת אחים למחצה היה צורך קודם בבדיקת הפגיעה בכל צמח (טבלה 2). נראה כי הצמחים הגדולים נפגעו בצורה דרסטית, ואילו הקטנים נפגעו פחות. לצאצאי T היה מקדם הרגרסיה השלילי ביותר, והם נזקקו למספר הימים הגבוה ביותר להתאוששות. לצאצאי G מקדם רגרסיית ההתאוששות הגבוה ביותר מבין כל שאר קבוצות הצאצאים (אך לא באופן מובהק), ואכן מספר ימי ההתאוששות שלו היה הנמוך ביותר. מקדם רגרסיית ההתאוששות של צאצאי GH,P ו-H אינו שונה מאפס, כלומר, אין עלייה בערכי הדיות במהלך ימי ההתאוששות. אחד ההסברים יכול להיות כי שלב ההתאוששות התרחש בחודש יולי, שיא הקיץ, ולכן גם לאחר החזרת ההשקיה המשיכו הצמחים בפעילות זהירה ושמרנית. לכן, בהתאוששות לא נמצאה עלייה בערכי הדיות לאורך הימים.
בבחינת ההתנהגויות השונות של כל צמח בכל שלב בניסוי על פי מוליכות הפיוניות (איור 2) נמצאו שתי אסטרטגיות התמודדות עם יובש המתאפיינות בשטח העלווה, באחוז איבוד העלים, ובסף סגירת הפיוניות ביובש:
ישנם צאצאים כדוגמת 8-1-GH שהם בעלי קצב גדילה אִטי אך בטוח, כיוון שהמוליכות נמוכה לאורך כל שעות היום ולאורך כל שלבי הניסוי, ומכאן הם מאופיינים בהתנהגות שמרנית איזוהידרית (איור 2ב). בצמחים אלה, כל העלים נשארו ירוקים וזקופים בסוף היובש (איור 2ב1), ומכאן המחשבה כי לא היה נזק הידראולי (בדיקות מוליכות הידראולית או קָוויטציה [cavitation] לא נעשו). העובדה ש-8-1-GH הראה התנהגות שמרנית במיוחד מבחינת מוליכות ואיבוד המים, יכולה להביא למסקנה כי היה שורד עוד ימים רבים ללא מים. נוסף על כך, הצמח 8-1-GH הוא הצאצא היחידי של העץ 8-GH בשנת 2013. מכאן עולה ההשערה כי עץ בתנאי מחיה קשים [4, 5], המייצר רק זרע אחד במשך עונת גידול שלמה, ייצר צאצא "שמרני" שיצליח לעמוד בתנאי יובש קשים מאוד. לתכונת השמרנות, כאמור, יתרונות לשמירת מאזן המים, אולם הצמח מסתכן בחוסר גדילה ובהרעבה לפחמן [19]. מעקב לאורך שנים יכול להביא לתשובה ברורה יותר: האם האסטרטגיה האיזוהידרית שבחרו 8-1-GH ודומיו להתחמקות מיובש, לא תפגע בהם גם לאחר שנים רבות? בהיבט אקולוגי, צמח המאופיין בגדילה אִטית בתנאים מיטביים, מסתכן בתחרות על אור ועל נוטריינטים עם העשבייה המקומית.
מהצד השני קיימים צאצאים כגון 10-KY, שהם בעלי שטח עלווה גדול ומוליכות פיוניות גבוהה, עם סף רגישות נמוך, כיוון שביובש איבד 100% מעליו (איור 2א). ייתכן שהפעילות הפיזיולוגית הנמרצת בתנאים מיטביים ובתנאי יובש הביאו ליתרון בזמן ההתאוששות, כיוון שהצמח הצליח להצמיח עלים חדשים (איור 2א1) ולהעלות מעט את המוליכות בשעות הבוקר (איור 2א). אמנם לא נעשו בדיקות קָוויטציה אך נראה כי הצמח "בוחר" לאבד את עליו למען מניעת קָוויטציה בגזע [29], וכניסה למצב "המתנה" עד שתכולת המים בקרקע תעלה. איבוד כל העלים מסכן את הצמח, כיוון שייתכן כי בתנאים טבעיים ההמתנה למים יכולה להימשך זמן רב. אולם, קצב הגדילה המהיר יעניק לצמח יתרון אקולוגי בתחרות על אור ואולי אף במרעה.
סיכום אופן הבחירה של עצי האֵם להעמדת צאצאים בתנאי סביבה משתנים מתואר באיור 3. עץ בתנאֵי מחיה גבוליים, סביר שיעמיד צאצאים שמרניים, כיוון שדור ההמשך כולו תלוי במספר קטן של צאצאים. עץ בתנאי מחיה טובים ייצר כמות בלוטים גדולה (עם שונות גנטית גדולה או קטנה) בעלי יתרון של צימוח מהיר. אין באסטרטגיה סיבה ותוצאה, אלא צירופי תכונות והתנהגויות, שיכולים להביא למסקנות עבור ייצור צאצאים עמידים ליובש. הכוונה, לדוגמה, אם צמח מגיע מעץ אֵם שנמצא בתנאי מחיה גבוליים, ונוסף על כך הצמח קטן ומתייבש לאט, ככל הנראה הוא יהיה בעל התנהגות איזוהידרית שמרנית, ומכאן – ישרוד יובש קיצוני במיוחד.
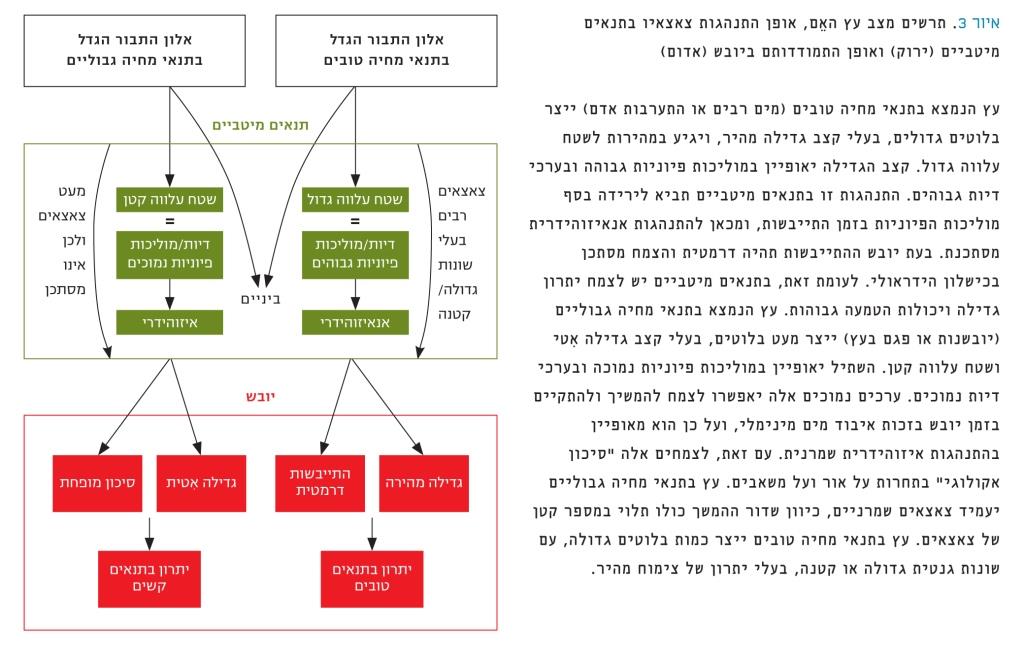
איור 3
תרשים מצב עץ האֵם, אופן התנהגות צאצאיו בתנאים מיטביים (ירוק) ואופן התמודדותם ביובש (אדום)
עץ הנמצא בתנאי מחיה טובים (מים רבים או התערבות אדם) ייצר בלוטים גדולים, בעלי קצב גדילה מהיר, ויגיע במהירות לשטח עלווה גדול. קצב הגדילה יאופיין במוליכות פיוניות גבוהה ובערכי דיות גבוהים. התנהגות זו בתנאים מיטביים תביא לירידה בסף מוליכות הפיוניות בזמן התייבשות, ומכאן להתנהגות אנאיזוהידרית מסתכנת. בעת יובש ההתייבשות תהיה דרמטית והצמח מסתכן בכישלון הידראולי. לעומת זאת, בתנאים מיטביים יש לצמח יתרון גדילה ויכולות הטמעה גבוהות. עץ הנמצא בתנאי מחיה גבוליים (יובשנות או פגם בעץ) ייצר מעט בלוטים, בעלי קצב גדילה אִטי ושטח עלווה קטן. השתיל יאופיין במוליכות פיוניות נמוכה ובערכי דיות נמוכים. ערכים נמוכים אלה יאפשרו לצמח להמשיך ולהתקיים בזמן יובש בזכות איבוד מים מינימלי, ועל כן הוא מאופיין בהתנהגות איזוהידרית שמרנית. עם זאת, לצמחים אלה "סיכון אקולוגי" בתחרות על אור ועל משאבים. עץ בתנאי מחיה גבוליים יעמיד צאצאים שמרניים, כיוון שדור ההמשך כולו תלוי במספר קטן של צאצאים. עץ בתנאי מחיה טובים ייצר כמות בלוטים גדולה, עם שונות גנטית גדולה או קטנה, בעלי יתרון של צימוח מהיר.
לסיכום, על פי המידע הקיים בידינו כעת, אנו ממליצים על צאצאי K, GH, P, KH כצאצאים עמידים ליובש קיצוני, ונראה כי לצאצאי K יתרון בעקבות אחוזי נביטה גבוהים (נספח 1) ובעלי קִרבה גנטית גדולה יחסית למרות האי-התאם העצמי (נספח 2ג). הצמחים הראו התנהגות שמרנית בטיפול היובש בניסוי זה. צמיחתם האִטית, אך הבטוחה, מקנה להם יכולת עמידות גבוהה ליובש על-ידי סף רגישות פיוניות גבוה (כשם שנמצא בעצי 'גבעת חציר'). לכן בזמן יובש, צמחים אלה לא איבדו כמעט את עליהם כיוון שככל הנראה לא היו בסכנה הידראולית. עקב מגבלת זמן נעצר ניסוי היובש לאחר 24 ימי יובש בלבד. מעניין היה לראות מה היה קורה אילו ניסוי היובש היה נמשך זמן רב יותר. ייתכן שצמחים אלה היו שורדים, אך במחיר גבוה של חוסר הטמעה לאורך תקופה ארוכה (הרעבה), ומכאן, שגדילתם האִטית של הצמחים מעמידה חיסרון בתחרות על אור ומשאבים. תוצאות המחקר מוגבלות, ודרוש מחקר מתמשך ורחב יותר כדי לקבוע בוודאות אילו עצים ישמשו מקור לזרעים. נוסף על כך, יש להתייחס למטען הגנטי המגיע מהעץ הזכרי (שאינו ידוע) ותורם גם הוא לתכונת העמידות.
תודות
תודה לקרן קיימת לישראל במימון המחקר ולאנשים שתרמו מזמנם: עינת גרזון, איילון כלב, אורי רוזנברג, אורי שמש, פרופ' יוסי ריוב, חברי המעבדה של ד"ר מנחם מושליון (זיו עטייה, גיל לרנר ונועם לוי), ד"ר ניר הר וחגי יבלוביץ' מקק"ל, פרופ' אמנון שוורץ וחברי מעבדתו (יותם זית ושראל מוניץ), ד"ר שבתאי כהן וויקטור ממנהל המחקר החקלאי – מרכז וולקני.
- התמודדות עם התייבשות עצים היא מאתגרי היערנות המרכזיים לנוכח שינוי האקלים והדיווחים על תמותת עצים ברחבי העולם. יש צורך מידי בגיבוש תכנית להשבחת עצים עמידים ליובש לצורך נטיעות.
- הבנת הבסיס הגנטי של אוכלוסיות עצים הכרחית להכנת תכנית שכזו, ובפרט כשמדובר בעצי יער שהידע הגנטי על אודותיהם מוגבל. התכנית יכולה לסייע בבחירת עצים שישמשו כמקור לזרעים.
- אלון התבור הוא מין מקומי וחשוב בייעור הארץ. מסתבר כי בין אלוני התבור הגדלים בבתי גידול שונים, ואף בין כאלה הגדלים באותו בית גידול, ישנם הבדלים ניכרים בהתנהגות הפיזיולוגית בתנאי יובש.
- המאמר מקדם ההבנה על אודות אופי הורשת תכונת העמידות ליובש באלוני התבור – תכונה חשובה לתכנון נטיעת יערות העתיד.
המערכת
נספחים
נספח 1. טבלת תיאור עצי האם ואחוזי נביטה
להורדהנספח 2א. טבלת רצף 11 ההתחלים ("פריימרים" F+R) וצבעם הפלורסנטי עבור ניסוי זה
להורדהנספח 2ב. עץ פילוגנטי של עצי האֶם וערכי ה- bootstrap
להורדהנספח 2ג. עץ פילוגנטי של עצי האֶם וצאצאיהם
להורדהנספח 3א. טבלת תוצאות גודל הסמנים בזוגות בסיסים (Allele) בעצי האֶם המשתתפים במחקר
להורדהנספח 3ב. טבלת תוצאות גודל הסמנים בזוגות בסיסים (Allele) עבור עצי האֶם (מודגש) וצאצאיהם, על פי ארבעת הסמנים הפולימורפים ביותר: ssrQpZAG9, ssrQpZAG15, ssrQpZAG36, ssrQpZAG119
להורדהמקורות
- אשד י. 1991. ריבוי אלונים מקומיים מבלוטים וייחורים (עבודת גמר לתואר מוסמך). רחובות: האוניברסיטה העברית בירושלים.
- גלון י. 2005. עצים בוגרים בישראל: סקר עצים בוגרים 1990–2005 וסיפורי עצים. משרד החקלאות ופיתוח הכפר.
- הלפרין ע. 2012. בחינת מנגנוני בקרה ותגובה של הצמח השלם לשינויים בזמינות המים ובדרישה האטמוספרית (עבודת גמר לתואר מוסמך). רחובות: האוניברסיטה העברית בירושלים.
- הר נ. 1998. מסלע וקרקע כגורם אקולוגי של תפוצה והתפתחות ביער אלון התבור באזור אלונים-שפרעם (עבודת גמר לתואר מוסמך). רחובות: האוניברסיטה העברית בירושלים.
- הר נ. 2008. מבנה מערכת הקרקע והסלע והדינמיקה של משק המים בבית הגידול כגורמים אקולוגיים עיקריים בתפוצת אלון התבור ואלון מצוי באזור אלונים-מנשה (חיבור לקבלת תואר דוקטור לפילוסופיה). רחובות: האוניברסיטה העברית בירושלים.
- זוהרי מ ופאהן א. 1981. צמחי התרבות של ישראל (מגדיר ומתאר). הקיבוץ המאוחד.
- קופר א. 2011. תנאי קרינה ביער מחטני נטוע והשפעתם על התפתחות ותפקוד אלון התבור. (עבודת גמר לתואר מוסמך). רחובות: האוניברסיטה העברית בירושלים.
- שילר ג. 2013. גידול עצי יער ויערות בישראל. ארץ של ספר המדבר.
- קליין ת, שפרינגר ע, פיקלר ב ואחרים. 2014. יעילות ניצול מים בעצי יער: עדיפות לאורן ירושלים על פני אלון מצוי. אקולוגיה וסביבה 5(3): 275–283.
- Almeida-Rodriguez AM, Cooke JE, Yeh F, and Zwiazek JJ. 2010. Functional characterization of drought-responsive aquaporins in Populus balsamifera and Populus simonii × balsamifera clones with different drought resistance strategies. Physiologia Plantarum 140(4): 321-333.
- Bréda N, Oszako T, and Delatour C. 2000. Water shortage as a key factor in the case of the oak dieback in the Harth Forest (Alsatian plain, France) as demonstrated by dendroecological and ecophysiological study. Recent advances on oak health in Europe; 22-24 Nov 1999; Warsaw, Poland.
- David TS, Henriques M, Kurz-Besson C, et al. 2007. Water-use strategies in two co-occurring Mediterranean evergreen oaks: Surviving the summer drought. Tree Physiology 27(6): 793-803.
- Dow B, Ashley M, and Howe H. 1995. Characterization of highly variable (GA/CT) n microsatellites in the bur oak, Quercus macrocarpa. Theoretical and applied genetics 91(1): 137-141.
- Ducousso A, Michaud H, and Lumaret R. 1993. Reproduction and gene flow in the genus Quercus L. Paper presented at the Annales des sciences forestières.
- Hagman M. 1975. Incompatibility in forest trees. Proceedings of the Royal Society of London, Series B. Biological Sciences 188(1092): 313-326.
- Kaplan D and Gutman M. 1999. Phenology of Quercus ithaburensis with emphasis on the effect of fire. Forest Ecology and Management 115(1):61-70.
- Lodhi MA, Ye GN, Weeden NF, and Reisch BI. 1994. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Molecular Biology Reporter 12(1): 6-13.
- Long TJ and Jones RH. 1996. Seedling growth strategies and seed size effects in fourteen oak species native to different soil moisture habitats. Trees 11(1): 1-8.
- McDowell NG, Pockman WT, Allen CD, et al. 2008. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist 178(4): 719-739.
- Ne’eman G. 1993. Variation in leaf phenology and habit in Quercus ithaburensis, a Mediterranean deciduous tree. Journal of Ecology 81: 627-634.
- Ne’eman G and Goubitz S. 2000. Phenology of east Mediterranean vegetation. In: Trabaud L (Ed). Life and Environment in the Mediterranean. Southampton, MA: Wit Press.
- Pou A, Medrano H, Tomàs M, et al. 2012. Anisohydric behaviour in grapevines results in better performance under moderate water stress and recovery than isohydric behaviour. Plant and Soil 359(1-2): 335-349.
- Sala A. 2009. Lack of direct evidence for the carbon-starvation hypothesis to explain drought-induced mortality in trees. Proceedings of the National Academy of Sciences of the United States of America 106(26): E68.
- Schiller G, Shklar G, and Korol L. 2003. Genetic diversity assessment by random amplified polymorphic DNA of oaks in Israel. 1. Tabor oak (Quercus aegilops L. ssp. ithaburensis [Decne] Boiss.). Israel Journal of Plant Sciences 51(1): 1-10.
- Schafale MP and Weakley AS. 1990. Classification of the natural communities of North Carolina. Third approximation. North Carolina Department of Environment, Health, and Natural Resources, Division of Parks and Recreation, Natural Heritage Program, Raleigh.
- Siwkcki R and Ufnalski K. 1998. Review of oak stand decline with special reference to the role of drought in Poland. European Journal of Forest Pathology 28(2): 99-112.
- Solomon S. 2007. Climate change 2007 – the physical science basis: Working group I contribution to the fourth assessment report of the IPCC 4: Cambridge University Press.
- Steinkellner H, Fluch S, Turetschek E, et al. 1997. Identification and characterization of (GA/CT) n-microsatellite loci from Quercus petraea. Plant molecular biology 33(6): 1093-1096.
- Tyree MT and Zimmermann MH. 2002. Xylem structure and the ascent of sap. Springer.
- Wallach R, Da-Costa N, Raviv M, and Moshelion M. 2010. Development of synchronized, autonomous, and self-regulated oscillations in transpiration rate of a whole tomato plant under water stress. Journal of Experimental Botany 61(12): 3439-3449.
- Zohary M. 1973. Geobotanical foundations of the Middle East: G. Fischer., Vol. 1,2, Stuttgart.




 מהדורה מודפסת
מהדורה מודפסת








