
חזית המחקר
יער או פער – התבססות אלון מצוי ביערות האורן בישראל
שני בן-יאיר בית הספר הבינלאומי ללימודי מדבר ע"ש אלברט כץ, אוניברסיטת בן-גוריון, קמפוס שדה בוקר
ברטרנד בוקן המחלקה לחקלאות באזורים צחיחים ע"ש ווילר, המכונים לחקר המדבר, אוניברסיטת בן-גוריון, קמפוס שדה בוקר
מאת
אורנע רייזמן-ברמן
המחלקה לחקלאות באזורים צחיחים ע"ש ווילר, המכונים לחקר המדבר, אוניברסיטת בן-גוריון, קמפוס שדה בוקרשני בן-יאיר
בית הספר הבינלאומי ללימודי מדבר ע"ש אלברט כץ, אוניברסיטת בן-גוריון, קמפוס שדה בוקרברטרנד בוקן
המחלקה לחקלאות באזורים צחיחים ע"ש ווילר, המכונים לחקר המדבר, אוניברסיטת בן-גוריון, קמפוס שדה בוקרמאת
אורנע רייזמן-ברמן
המחלקה לחקלאות באזורים צחיחים ע"ש ווילר, המכונים לחקר המדבר, אוניברסיטת בן-גוריון, קמפוס שדה בוקרשני בן-יאיר
בית הספר הבינלאומי ללימודי מדבר ע"ש אלברט כץ, אוניברסיטת בן-גוריון, קמפוס שדה בוקרברטרנד בוקן
המחלקה לחקלאות באזורים צחיחים ע"ש ווילר, המכונים לחקר המדבר, אוניברסיטת בן-גוריון, קמפוס שדה בוקרהדור הראשון של יערות המחט בישראל מתחלף כיום בדור שני – יער מגוון המורכב גם ממיני חורש מעוצים רחבי עלים. אחת השאלות המרכזיות היא, האם ובאילו תנאי סביבה יכולים מיני חורש רחבי עלים להתבסס ביערות האורן.
מאמר זה מעלה היבטים אקולוגיים, פיסיולוגיים ויערניים, הנוגעים לשאלה זו, אשר עשויים להוות בסיס להעלאת השערות בנוגע לתהליכי ההתבססות של מיני חורש ביער האורן. המאמר מציג תוצאות ראשוניות של מערכת ניסויית העוסקת בחלק מן ההיבטים של שאלה זו, במין אלון מצוי – המין הדומיננטי בחורש הים-תיכוני בארצנו. בניסוי נבחנו השפעת הגורמים הבאים על התבססות האלון ביער האורן: (א) מידת פתיחות החופה הנקבעת על ידי מבני יער שונים: פער, שולי פער וצפיפויות משתנות של עצי היער. כמו כן, הניסוי בוחן את השפעת מידת זמינות האור לאורך גרדיאנט של הצללה מלאכותית; (ב) מפנה; (ג) שונות בין שנים. השערת הניסוי הייתה, שסביבת היער עשויה להיטיב עם ההתבססות של האלון המצוי בשל ההקלה הנובעת ממנה בתנאי שההצללה לא תהיה כבדה מדי. תוצאות הניסוי מלמדות, כי הצללה קלה ובינונית, בין שהיא מושרית על ידי היער ובין שהיא מושרית על ידי רשתות צל, אכן משפרת את שיעור ההתבססות, אולם התפתחות האלון בפער נמרצת וטובה יותר. המאמר דן בניגודיות זו ובהשלכותיה על ממשק היער.
מבוא
יערות האורן בישראל הם נוף תרבותי ומערכת אקולוגית ייחודית. את דור היער הראשון, המורכב בעיקר ממחטניים, מחליף כיום דור היער השני, הוא "היער החדש" – יער מגוון המורכב גם ממיני חורש מעוצים רחבי עלים [22]. תהליך היווצרות היער החדש הוא תולדה של ממשק, דינמיקה טבעית והפרעות, בעיקר שרפות.
מגמת התהליך, המסמנת את היווצרותו של יער מעורב (mixed forest), מעלה שאלות הנוגעות לאפיון הרכב היער (composition) ומבנה היער (structure) העתידי, ליכולתנו לחזות את אופיו של היער ואת הדינמיקה שלו, וכן שאלות הנוגעות להתאמת ממשק ליצירת יער מעורב בר-קיימא.

כיום, אפשר למצוא מגוון רב של "יערות חדשים" במצבים שונים, החל ביערות אשר בהם רחבי העלים מהווים תת-יער תחת חופת היער החוסה וכלה ביערות מתחדשים שבהם רחבי העלים מהווים מרכיב עיקרי. עם זאת, יש גם יערות שבהם התבססות רחבי העלים היא אפסית או מועטה ביותר.
אחת השאלות המרכזיות היא, האם ובאילו תנאי סביבה יכולים מיני חורש רחבי עלים להתבסס ביערות האורן.
מקובל להניח, שתהליכי ההתבססות של אוכלוסיות צמחים תלויים בשני גורמים עיקריים: (א) זמינות הזרעים באתר; (ב) זמינות מיקרו האתרים אשר התנאים בהם מתאימים להתבססות [11, 20, 34]. מאמר זה מתמקד בתהליכי ההתבססות, בהנחה שכמות הזרעים אינה מוגבלת. המאמר מתרכז בהיבט האקו-פיזיולוגי, בהיבט העתי-מרחבי ובהיבט המקומי של ההתבססות במערכות יער באזור ים-תיכוני, המאופיין בעקות יובש הן בקיץ והן בעונת הצמיחה.
באזורי אקלים, כמו האזור הים-תיכוני היובשני, חופת היער יוצרת סביבה מוצלת המאופיינת בקרינה מופחתת, בטמפרטורות נמוכות ובלחות גבוהה יותר בהשוואה לסביבה היובשנית ללא חופה. למרות ההצללה, הסביבה יובשנית וחשופה לעקות יובש. לפיכך, התבססות והתפתחות מוצלחת של צומח תחת חופה דורשת התמודדות בו-זמנית עם הצללה ועם עקת יובש.
כיום, ישנן שתי השערות מרכזיות המציגות מגמות מנוגדות בנוגע להשפעת ההצללה, המשולבת עם עקת יובש, על התבססות מיני צמחים והתפתחותם. ההשערה הראשונה היא "השערת יחסי ההקלה" [8] (facilitation). על פי השערה זו, התנאים הנוצרים תחת חופת הצומח באזורים אשר בהם שוררות עקות (למשל עקת יובש או קור), עשויים להקל על התבססותם של מיני צומח שונים. על כן, עם העלייה בחומרת העקות הסביבתיות, תימצא עלייה במקרי "יחסי ההקלה". ואכן, יש מחקרים המורים כי באזורים יובשניים, התבססות צמחים "בני-חסות" תחת חופת הצמח "החוסה" היא הגורם המרכזי הקובע את דינמיקת הצומח באזורים אלה [14].
ההשערה האחרת היא השערתם רבת ההשפעה של [29] Smith and Huston, המכונה "השערת ההמרות" (trade-off hypothesis). על פי השערה זו, צמחים אינם מסוגלים להתמודד בו-זמנית עם עקת מים ועם הצללה, משום שהמנגנונים הפיזיולוגיים והמורפולוגיים המאפשרים התמודדות עם הצללה נוגדים את המנגנונים המאפשרים התמודדות עם עקת יובש. לדוגמה, מנגנון הגדלת שטח העלווה, שהינו מנגנון להתמודדות עם הצללה, נוגד את מנגנון הקטנת שטח העלווה, הפועל בתנאי עקת מים. אם השערה זו נכונה, הרי שלמעשה רק מיני צמחים מעטים יכולים להתבסס בתנאים של יובש והצללה, ומגוון האתרים שבהם ההתבססות אפשרית בכלל – מצומצם.
מחקרים במספר רב של מיני מעוצים, בעיקר מהאזורים הממוזגים [21] מלמדים, כי היכולת להתמודד בו-זמנית עם כמה עקות – נדירה. ישנם מעט מאוד מחקרים אשר נערכו במיני מעוצים ים-תיכוניים. Sanchez-Gomez ועמיתיו [28] בחנו את שרידות הנבטים במיני מעוצים ים-תיכוניים צפוניים והראו, כי יכולת התמודדות בו-זמנית עם עקת צל ועם עקת יובש היא תלויית-מין (species-specific), כלומר, חלק מן המינים עמידים לשתי העקות בו-זמנית וחלקם אינם עמידים. במינים ים-תיכוניים יובשניים יותר, כגון Q. ilex, Quercus coccifera מינים הדומים לאלון המצוי (Q. calliprinos) ואלת המסטיק (Pistacia lentiscus) נמצא, כי ההשפעה המעכבת של עקת היובש הן על שרידות הנבטים והן על התפתחותם הייתה חמורה יותר בחשיפה לקרינה מלאה לעומת השפעתה תחת הצללה [27]. לעומת זאת, Maestre ועמיתיו [18] בחנו את התבססותם של מינים אלה ועוד כמה נוספים ביערות האורן בספרד ומצאו, כי חופת היער אינה מקלה על ההתבססות ואינה משפרת את תפקודם הפיזיולוגי בהשוואה להתבססות בקרחות יער. במיני מעוצים ים-תיכוניים אחרים נמצא, כי ההצללה אף מחריפה את השפעת הבצורות וגורמת לירידה משמעותית בתפקוד הפיזיולוגי ובשיעור הישרדות הנבטים [18, 31, 32]. ההשפעות המיטיבות או המעכבות של ההצללה עשויות להשתנות גם עם עונות השנה [33]. עוצמת ההצללה קובעת גם היא את מידת ההטבה. כך למשל, עוצמת הצללה בינונית עשויה לשפר את ההתבססות אפילו במיני חלוץ דורשי אור כמו סירה קוצנית, ואילו הצללה כבדה מעכבת את ההתבססות [24]. נמצא גם, כי ההטבה הניתנת על ידי ההצללה תלויה בשלב ההתבססות או ההתפתחות של "בן החסות", כך ששלבי ההתבססות הראשוניים דורשים צל, אולם ההתפתחות וההתבססות ארוכת הטווח דורשת חשיפה לקרינה מלאה [19 ,10].
השאלה בדבר השפעת ההצללה על התבססות והתפתחות של צומח בתנאים יובשניים נותרת בלתי פתורה. אם כך, אפשר להעלות מספר השערות חלופיות על מידת הצלחת ההתבססות של מיני החורש ביערות האורן בארץ.
הדינמיקה הטבעית של אוכלוסיות וחברות של מיני עצים יוצרת ביער שונוּת במרחב ובזמן. השונות עשויה לבוא לידי ביטוי במגוון מיני העצים, גילם ופיזורם במרחב היער. היער מאופיין בכתמי יער שונים כפערי יער (קרחות יער ,forest gaps) וכתמים בעלי צפיפויות עצים משתנות. כתמי היער נבדלים אלה מאלה במיכלול של תנאים סביבתיים המושפעים ממידת פתיחות החופה בכתם, כמו למשל, עוצמת הקרינה, הטמפרטורה, לחות הקרקע ומשטר העירבול של זרמי האוויר [12]. הדבר נכון הן ליערות טבעיים והן ליערות נטע אדם, הניטעים בצפיפויות שונות והעוברים תהליכי דילול יערני או טבעי.
הפערים ביער נוצרים בשל תמותת עצים בוגרים או בגלל הפרעה. תהליכי היווצרות הפערים ואכלוסם המחודש ידועים בשם "דינמיקת הפערים" [15] (gap dynamics). על-פי השערת הסוקצסיה של הפער או השערת ההתמיינות לנישות (niche partitioning), פערי יער מתאכלסים בהתאם לדרישות המינים למשאבים שונים, ובעיקר הדרישה לאור [25]. השערה חדשנית יותר טוענת כי התבססות המינים בפער היא אקראית וקשה לחיזוי [9]. דינמיקת הפערים היא התהליך המרכזי היוצר את פסיפס הכתמיות המרחבית והקובע את מגוון מיני היער [30]. העקרונות האקולוגיים של דינמיקת הפער מיושמים בממשק היער. לדוגמה, נעשה שימוש ביצירת פערים מלאכותיים לצורך עידוד ההתבססות של מינים חדשים הדורשים תנאי הארה טובים יותר [7].
בארץ ישנו מידע מועט בנושא תרומת הפער וכתמי יער בצפיפויות שונות למבנה היער ולדינמיקה שלו. בשל כך, קשה לחזות את יכולת ההתבססות וההתפתחות של מיני החורש ביער המחטני. כמו כן, עד כה בוצעו מספר מצומצם של מחקרים על השפעות ההצללה בכלל, והצללת יער בפרט, על מיני חורש מקומיים. מחקריו של חריף (1974) באלון מצוי הראו, כי התבססותו בחורש מוצלחת יותר באתרים מוצלים לעומת אתרים החשופים לקרינה [3]. גם מחקרו של גולדרינג (1977) הראה, כי שרידות נבטי האלון המצוי הייתה הגבוהה ביותר בהצללה כבדה אשר מושרית ביער הוותיק [2].
מחקרם של קדמון ואלון (1996) מעלה, כי התבססות נבטי אלון מצוי מבלוטים הייתה הגבוהה ביותר תחת שיחי סירה קוצנית. זאת, בהשוואה לקרחות יער, לקרחות בתה ולחורש צפוף [6]. כלומר, הצללה שאינה כבדה מדי עשויה להיטיב עם שרידות האלון. תוצאות ניסוי זה מחזקות את הצורך לבחון את מידת ההצללה העשויה להיטיב עם ההישרדות ועם ההתפתחות של מיני חורש רחבי עלים מקומיים.
מחקרים על התפתחות האלון המצוי בתנאים מבוקרים מלמדים, כי תחת הצללה מערכת השורשים של האלון מתפתחת מעט ביחס להתפתחות הנוף העל-קרקעי [1], תופעה המוכרת כתגובה לצל [16]. טפר (1997) בחן את התפתחותם של שתילי אלון מצוי תחת משטרי מים והארה שונים ומצא, כי כאשר זמינות המים נמוכה יחסית, התפתחות האלון בתנאי הארה מלאה הייתה טובה יותר בהשוואה להתפתחותו בתנאי הצללה [4].
כדי לבחון כמה מן ההיבטים המובאים לעיל חקרנו את יכולת ההתבססות של המין אלון מצוי ביער אורן. מין זה נבחר בשל היותו המין המעוצה הדומיננטי בחורש הים-תיכוני בארצנו. נבחנה השפעת: (א) מידת פתיחות החופה הנקבעת על ידי מבני יער שונים: פער, שולי פער וצפיפויות משתנות של עצי היער. כמו כן, נבחנה השפעת מידת זמינות האור לאורך גרדיאנט של הצללה מלאכותית. זאת, כדי להבין את השפעת הירידה בעוצמת ההארה לעומת השפעת היער כמיכלול; (ב) מפנה; (ג) שונות בין שנים. השערת הניסוי הייתה, שסביבת היער עשויה להיטיב עם ההתבססות של האלון המצוי, בתנאי שההצללה לא תהיה כבדה מדי. השערה נוספת היא, כי ההטבה תהיה משמעותית יותר במפנה הדרומי בהשוואה למפנה הצפוני.
שיטות
אתר הניסוי: הניסוי נערך ביער משואה (619458 / 192712 ITM). האקלים הוא ים-תיכוני יובשני (כ-400 מ"מ ממוצע משקעים שנתי). הקרקע היא רנדזינה בהירה על גבי קירטון איאוקני [5]. נבחרו עומדים של אורן ירושלים (Pinus halopensis) אשר ניטעו בסוף שנות ה-1950 או בתחילת שנות ה-1960.
מבנה הניסוי: מבנה הניסוי מקונן וכולל 12 יחידות של פער מלאכותי (הנוצר בכריתה) ואת היער בסביבתו, על גבי מפנים צפוניים ודרומיים. במפנה הדרומי של אתר הניסוי צפיפות העצים הייתה גבוהה (כ-100 עצים לדונם), לכן דולל היער ליד הפער לכ-40 עצים לדונם – צפיפות המאפיינת את המפנה הצפוני. באופן זה יצרנו גרדיאנט של פתיחות, חופה והצללה, הכולל את: (א) הפער אשר בו אזורים החשופים לקרינה רוב שעות היום; (ב) שולי הפער, החשופים לקרינה רק בחלק משעות היום, בבוקר או אחר הצהריים; (ג) יער דליל; (ד) יער צפוף (רק במפנה הדרומי). בכל יחידת פער הוצבו חלקות (1.5 מטר X 1.5 מטר). בנוסף, הוצבו בפער חלקות מוצללות ברשת צל (50, 70 ו-90 אחוזי הצללה), כך שבכל יחידת פער עד 7 חלקות טיפול. בכל חלקה נזרעו בחורף 2006/07 64 בלוטים אשר נאספו באזור. החלקות מוגנות מטריפה בעזרת רשת לול.
גודל הפערים תוכנן על ידי מודל ממוחשב, ועל פי הטופוגרפיה של יער משואה, כך שגודל הפער יאפשר למקם ארבע חלקות החשופות במשך כל השנה להארה לכל הפחות בשעות מ-10:00 ועד ל-15:00, ביום הקצר ביותר. על סמך המודל נקבע גודל הפער ל-15 מטר X 25 מטר. כדי לבחון את השפעת השונות בין שנים על ההתבססות, נזרעו בכל חלקה 20 בלוטים נוספים, בחורף 2007/08.
תצפיות: מעקב אחר השרידות וההתפתחות נערך אחת לשישה שבועות בממוצע. מדדי ההתפתחות נמדדו במדגם של שמונה נבטים לחלקה, והם כללו קוטר בסיס, גובה, מספר העלים ומספר ההסתעפויות מהגבעול המרכזי.
חישובי שיעורי ההתבססות: שיעורי ההתבססות מחושבים כמכפלה שבין שיעורי ההצצה לשיעורי השרידות. שיעורי ההצצה הם מספר המציצים מתוך 64 הבלוטים שנזרעו, ושיעורי השרידות הם מספר השורדים מתוך מספר המציצים. שיעורי התמותה חושבו על סמך השיפוע של קצב השרידה, כלומר, מספר השורדים במועד מסוים פחות מספר השורדים במועד מאוחר יותר, מחולק במשך הזמן שעבר בין המועדים.
ניתוח סטטיסטי: ניתוח השונות נערך במודל מקונן (פער מקונן במפנה), שבו נבחנו השפעת המפנה, ההצללה והאינטראקציה ביניהם, כהשפעות עיקריות. השוואות בין טיפולים נערכו במבחני Tukey’s HSD pairwise comparisons או ב-contrasts. תהליך ההתבססות נבחן ב-repeated measures לאורך זמן, כאשר בנוסף להשפעות של הטיפולים העיקריים נבחנה גם השפעת הזמן והאינטראקציה שלה עם טיפולי המפנה וההארה. הניתוח נערך בתוכנה Statistica.

תוצאות
תהליכי הצצה, הישרדות והתבססות:
הצצת הנבטים מהבלוטים אשר נטמנו בדצמבר 2006 הייתה דומה בכל טיפולי ההצללה (p > 0.05), ושיעורה היה 67±1% בממוצע, מתוך כלל הבלוטים שהוטמנו. הן להצללות השונות והן למפנה הייתה השפעה מובהקת על שיעורי ההתבססות (p < 0.001). השפעת טיפולי ההצללה השתנתה עם הזמן במהלך ההתבססות (0.01 >p) וכך גם השפעת המפנה (0.0001 >p) (איור 1). השינוי בהשפעת המפנה וההצללה עם הזמן ניכר בכך, שבשלבי ההתבססות הראשוניים לא נמצאו הבדלים מובהקים בשיעורי ההצצה וההתבססות בין שילובי הטיפולים השונים. אולם, באפריל 2007 החלה תמותת הנבטים, כאשר התמותה החריפה ביותר חלה במפנה הדרומי בפער ובשולי הפער (איור 1). לעומת זאת, התמותה במפנה הצפוני תחת הצללת רשת של 50 ו-70 אחוזים הייתה הנמוכה ביותר. כמו כן, במפנה הדרומי חלה תמותה גבוהה יותר לאחר העונה היבשה השנייה (קיץ 2008), ואילו במפנה הצפוני כמעט שלא הייתה תמותה. בניתוח שיעורי התמותה נמצא, כי בשנת ההתבססות הראשונה (אפריל 2007 עד אפריל 2008) הייתה הן להצללה והן למפנה השפעה מובהקת על שיעורי התמותה (p < 0.01) ואילו החל מאפריל 2008 הייתה למפנה בלבד השפעה מובהקת על התמותה (p < 0.01).
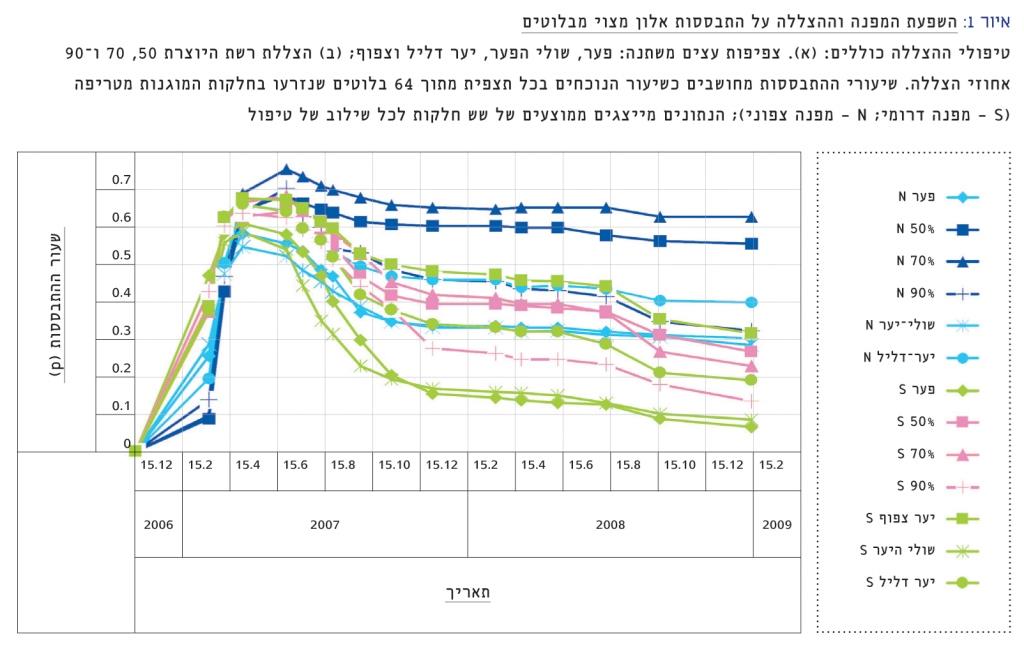
איור 1
השפעת המפנה וההצללה על התבססות אלון מצוי מבלוטים
טיפולי ההצללה כוללים: (א). צפיפות עצים משתנה: פער, שולי הפער, יער דליל וצפוף; (ב) הצללת רשת היוצרת 50, 70 ו-90 אחוזי הצללה. שיעורי ההתבססות מחושבים כשיעור הנוכחים בכל תצפית מתוך 64 בלוטים שנזרעו בחלקות המוגנות מטריפה (S – מפנה דרומי; N – מפנה צפוני); הנתונים מייצגים ממוצעים של שש חלקות לכל שילוב של טיפול
לאחר 26 חודשים ממועד הזריעה היה שיעור ההתבססות הממוצע במפנה הצפוני גבוה פי שניים באופן מובהק (p < 0.001) משיעור התבססות הממוצע במפנה הדרומי (40±5% לעומת 20±3%, בהתאמה). בין טיפולי ההצללה, שיעורי ההתבססות הנמוכים ביותר היו בפער ובשולי הפער ואלה היו שונים באופן מובהק (Tukey’s HSD test p < 0.05) מהשיעורים הגבוהים ביותר אשר נצפו בהצללה של 50 ו-70 אחוזים.
שיעורי ההישרדות בפער (שורדים מתוך סך כל המציצים) לאחר 26 חודשים מן הזריעה, היו נמוכים באופן מובהק משיעורי ההישרדות ביער ככלל (דליל וצפוף, בשני המפנים כאחד – p < 0.05 contrasts).
רצוי לשים לב, שבניגוד להטבה שיצרו ההצללות המלאכותיות של 50 ו-70 אחוזים, שיעורי ההתבססות תחת רשת ההצללה של 90% היו נמוכים יחסית ודומים לשיעורי ההתבססות בפער ובשולי הפער.
המגמה המסתמנת היא, כי ההתבססות במפנה הצפוני גבוהה מזו שבדרומי, ואילו ההצללה, בין אם היא מושרית באופן מלאכותי ובין אם היא מושרית על ידי חופת היער, מיטיבה עם ההתבססות והשרידות של האלון המצוי. הצללת רשת משפרת עד מאוד את ההתבססות במידה שאינה כבדה מדי (90%). הצללה כבדה מעכבת את ההתבססות.
התפתחות נבטי האלון:
קוטר הבסיס של פרטים בני 26 חודשים הושפע באופן מובהק מההצללה (p < 0.000001), אך לא מהמפנה. קוטר הבסיס של פרטים אשר צמחו בפער היה הגבוה ביותר באופן מובהק (Tukey’s HSD test p < 0.05) ואילו פרטים אשר צמחו בתנאי היער הצפוף ובתנאי הצללת רשת של 90% היו בעלי הקוטר הנמוך ביותר, בכ-30% נמוך מצמחי הפער (איור 2). מבין תנאי ההצללה, הפרטים אשר צמחו תחת חופת היער היו בעלי קוטר נמוך מאלה שצמחו תחת רשתות הצללה של 50 ו-70 אחוזים. גובה הפרטים גם הוא הושפע באופן מובהק מטיפולי ההצללה (p< 0.000001), אך לא מן המפנה. כצפוי, פרטים אשר צמחו בהצללת הרשת היו הגבוהים ביותר באופן מובהק, ב-30% יותר מאלה שבהארה מלאה בפער (איור 2). שלא כצפוי, פרטים אשר צמחו תחת חופת היער היו הנמוכים ביותר באופן מובהק, נמוכים אף מאלה שגדלו בפער (איור 2). ההצללה השפיעה באופן מובהק גם על מספר ההסתעפויות מן הגבעול המרכזי (p< 0.000001) ועל מספר העלים (p< 0.000001). פרטים אשר צמחו בפער היו גם בעלי מספר ההסתעפויות הגבוה ביותר (2.6 ± 0.2) וגם בעלי מספר העלים הגבוה ביותר (126 ±), ואילו הפרטים שצמחו ביער הדליל, הצפוף, ובהצללה של 90%, היו כמעט מחוסרי הסתעפויות ומספר העלים היה כ-10. התוצאות מלמדות, כי תנאי הפער מיטיבים עם התפתחות האלון המצוי. לעומת זאת, תנאי היער מעכבים את התפתחות האלון גם בהשוואה לתנאי הצללה קלה ובינונית על ידי רשת צל. הצללה כבדה (90%) על ידי רשת צל מעכבת גם היא את ההתפתחות.
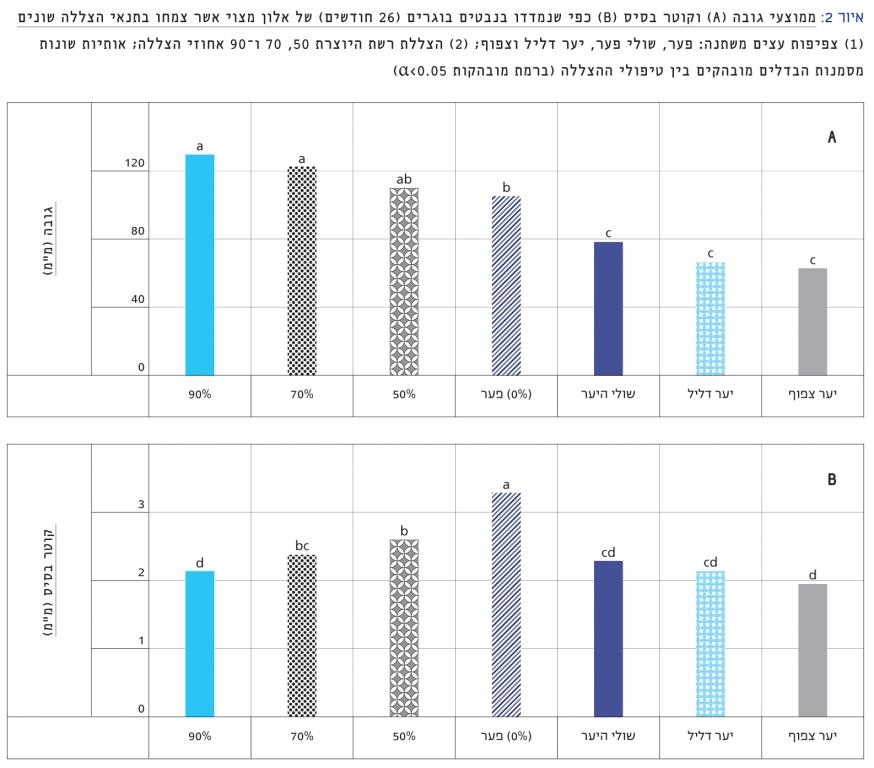
איור 2
ממוצעי גובה (A) וקוטר בסיס (B) כפי שנמדדו בנבטים בוגרים (26 חודשים) של אלון מצוי אשר צמחו בתנאי הצללה שונים
(1) צפיפות עצים משתנה: פער, שולי פער, יער דליל וצפוף; (2) הצללת רשת היוצרת 50, 70 ו-90 אחוזי הצללה; אותיות שונות מסמנות הבדלים מובהקים בין טיפולי ההצללה (ברמת מובהקות 0.05<α)
השפעת השונות בין שנים על תהליכי ההתבססות:
לשנה שבה נטמנו הבלוטים (דצמבר 2006 או דצמבר 2007) הייתה השפעה מובהקת על שיעורי הצצת נבטי האלון (p < 0.01), כפי שנצפו כשלושה חודשים לאחר ההטמנה (2007 ו-2008 בהתאמה). כמו כן, לשנת הזריעה הייתה השפעה מובהקת גם על שרידות הנבטים (p < 0.000001) וגם על התבססותם (p < 0.000001). התוצאות מורות על התבססות גבוהה פי שתיים בחורף 2006/07 (איור 3). בהשוואה בין שיעורי השרידות וההתבססות של שני הגייסות (cohort) של 2007 ו-2008, שמונה חודשים לאחר ההטמנה של כל גייס, נמצא כי שני הגייסות הושפעו באופן דומה מן ההצללה, אך השפעת המפנה הייתה שונה בין הגייסות (מפנה X שנה, שרידות p < 0.01, התבססות p < 0.05). בגייס 2007 לא הייתה למפנה כל השפעה על שיעורי ההתבססות, כפי שנצפו שמונה חודשים לאחר ההטמנה, ואילו בגייס 2008 הייתה השפעת המפנה מובהקת (p < 0.01) ושיעורי ההתבססות במפנה הדרומי היו נמוכים פי שניים משיעורי ההתבססות במפנה הצפוני (איור 3). בבחינת השפעת ההצללה על פני שתי השנים נמצא, כי שיעורי ההתבססות בפער ובשולי הפער הם הנמוכים ביותר באופן מובהק (Tukey’s HSD test p < 0.05) ושיעורי ההתבססות תחת רשתות ההצללה הם הגבוהים ביותר באופן מובהק. שיעורי ההתבססות ביער גבוהים באופן מובהק מאלה שבפער ובשולי הפער. המגמות הדומות בשתי השנים בדבר השפעת תנאי ההצללה מחזקות את ההשפעה המיטיבה שיש ליער ולהצללה מלאכותית על שלבי ההתבססות הראשוניים.

איור 3
שיעורי ההתבססות של נבטי אלון מצוי בני שמונה חודשים, אשר נזרעו בשתי שנים שונות (דצמבר 2006 ודצמבר 2007)
האיור מציג את ממוצעי ההסתברות (+ שגיאת תקן) להתבססות מוצלחת של שני הגייסות בכל אחד משילובי הטיפולים שמונה חודשים לאחר הזריעה; הבלוטים נזרעו בחלקות מוגנות מטריפה על גבי מפנה צפוני ודרומי; טיפולי ההצללה כוללים: (א) צפיפות עצים משתנה: פער, שולי פער, יער דליל וצפוף; (ב) הצללת רשת היוצרת 50 ,70 ו-90 אחוזי הצללה; שיעורי ההתבססות מחושבים כשיעור הנוכחים מתוך הבלוטים שנזרעו
דיון
ממצאי ניסוי זה מראים, כי לשלבים שונים בהתבססותו של האלון המצוי דרושים תנאי סביבה שונים. צל עצי היער (מלבד בשולי הפער) וכן ההצללה המלאכותית היטיבו עם שלבי ההתבססות הראשוניים של האלון ושיפרו את הישרדות האוכלוסייה בהשוואה למידת הישרדותה בפער, בהארה מלאה. אולם, התפתחות הפרטים בפער בחשיפה להארה מלאה הייתה טובה לאין ערוך מזו שבתנאי ההצללה. למעשה, מתגלה כאן ניגודיות בין הישרדות להתפתחות: בפער השרידות נמוכה, אך ההתפתחות מואצת, וביער השרידות גבוהה, אך ההתפתחות מעוכבת או אפילו מדוכאת.
בנוסף לכך, התוצאות מצביעות על האפשרות, שהאלון המצוי אינו יכול להתמודד עם הצללה כבדה. לעומת זאת, הצללה בינונית משפרת עד מאוד את השרידות בתנאי היובש הים-תיכוניים. מגמה זו תואמת את המסגרת הרעיונית אשר העלו Holmgren ועמיתיה [17] בניסיון לפתור את הסתירה הקיימת בין "השערת ההמרות" של [29] Smith and Huston לבין התצפית המורה, כי דווקא באזורים יובשניים ישנה התבססות גבוהה בצל חופת הצומח. החוקרים טענו, כי הצל יכול להקל את עקת היובש ולשפר את תפקוד הצמח, בעיקר באמצעות שמירה על מאזן המים, כל עוד זמינות האור אינה הופכת לגורם המגביל.
השוואה בין התבססות מינים שונים של שיחים ממשפחת הסלקיים באזור יובשני באוסטרליה הראתה, כי מידת הסבילות (tolerance) לצל היא המשפיעה על היעילות ועל טווח ההקלה הניתנת על ידי הצמח החוסה [23]. תופעת הסבילות לצל נמצאה גם במחקר זה. צמחי האלון אשר צמחו תחת רשתות ההצללה פיתחו מורפולוגיה של "הימנעות מצל" (shade avoidance). מורפולוגיה זו מאופיינת בצימוח נמרץ של הגבעול המרכזי ללא הסתעפויות או עם מעט הסתעפויות. אופן התפתחות זה מאפשר צימוח לזמן מה בסביבה מוצלת, התחמקות ממנה בעזרת התארכות הגבעול והתפתחות מאוחרת יותר באור [16], כמו למשל, במקרים שבהם ישנה התבססות מתחת לשיחים. אפשר לומר, כי רוב מיני המעוצים הים-תיכוניים של האזורים היובשניים הם דורשי אור ויכולים לסבול צל לתקופה קצרה במחזור חייהם [33].
בניגוד לפרטים אשר צמחו תחת רשתות ההצללה, אלה שצמחו בפער היו בעלי נוף מפותח, מסועף ומרובה עלים. הפרטים אשר צמחו ביער, לעומת זאת, היו מפותחים פחות מאלה שצמחו תחת רשתות הצל. עוצמת ההארה ביער היא כ-60% מעוצמת ההארה בפער (נתונים לא מובאים) ולמרות זאת, התפתחות הפרטים אשר צמחו ביער ובשולי הפער לא דמתה להצללה של 50%. ניתן להעלות כמה השערות בנוגע להתפתחות הלקויה. אפשרות אחת היא, כי העיכוב בהתפתחות נובע מספקטרום ההארה מתחת לחופת היער, אשר הינו שונה מהספקטרום בפער (רשתות הצל אינן משנות את הספקטרום אלא מפחיתות את עוצמת הקרינה). תופעות מעין אלו נצפו במינים שאינם סבילים לצל [13]. אפשרות אחרת היא תחרות עם העצים הבוגרים ביער, אולי בעיקר תחרות על משאב המים. במדידות שערכנו בעומקים של עד 40 ס"מ לא מצאנו הבדלים מובהקים בלחות הקרקע בין היער לפער, וככלל לא נמצאו הבדלים מובהקים בין כל הטיפולים (נתונים לא מובאים).
הניגודיות שבין השרידות להתפתחות מעלה את השאלה – כיצד יכולה להתפתח אוכלוסיית אלונים מבוססת? האם יש חשיבות גדולה יותר למספר הפרטים או למידת התפתחותם?
ביערות באזור הממוזג נצפה שמעוצים מנוונים המכונים "גמדי צל", יכולים לשרוד במצב זה לאורך שנים והם מתפתחים רק במידה והם נחשפים לאור [16, 30]. לעומת זאת, בדומה למחקר זה, הראו צמחי אלון מתבססים של Q. petraea ו-Q. pyrenaica התפתחות לקוייה, תיפקוד ירוד, ועמידות נמוכה ליובש, בתנאי יער הנטוע בצפיפות גבוהה לעומת בינונית. זאת, למרות שלחות הקרקע הייתה דומה בשני הטיפולים [26]. יותר מכל, ייתכן שהאלון המצוי זקוק להצללה רק בשלבים הראשונים של ההתבססות, וחשיפה לאור נחוצה לצורך התפתחות מלאה, כפי שנמצא במינים אחרים של אלון [10]. מבחינת התבססות האוכלוסייה, עולה כאן האפשרות שעל אף השרידות הנמוכה בפער, ייתכן שההתפתחות הטובה בפער עשויה לתמוך בהתבססות מהירה יותר של אוכלוסיית האלון. כך למשל, צמחים אשר צומחים בפער יכולים להגיע לשלב עשיית הבלוטים מהר יותר וייתכן שפרטים בודדים יוכלו לבסס אוכלוסייה במהירות רבה יותר. תפקיד הפער בביסוס האוכלוסייה מתחזק לנוכח התוצאות של ניסוי מקדים במחקר זה, אשר בחן את טריפת הבלוטים. נמצא, כי שיעור טריפת הבלוטים גבוה ביער ונמוך מאוד עד אפסי במרכז הפער. מכאן ומתוצאות המחקר המוצגות אפשר ללמוד, כי ההתבססות תלויה במספר רב של גורמים, שקיימת ביניהם אינטראקציה. ממשק יער מעורב דורש אפוא שקלול של כמה גורמים, אשר בחלקם פועלים בכיוונים הפוכים.
ממצאים נוספים ממחקר זה מלמדים, כי נוסף על השפעת ההצללה, למפנה ולשונות בין שנים הקשורה בכמות המשקעים יש השפעה ניכרת על שיעורי ההתבססות. ההתבססות במפנה הדרומי נמוכה יותר, ובפער הדרומי שיעורי ההתבססות נמצאו נמוכים עד מאוד. בשנת בצורת (חורף 2007/2008) נמצאה השפעה מובהקת של המפנה גם על תהליך ההצצה, שהייתה נמוכה יותר במפנה הדרומי. תופעות אלו מוכרות ותומכות במחקרים קודמים באזורים ים-תיכוניים יובשניים [35]. אולם, מחקר זה מציג ממצא מעניין חדש המורה, כי למרות השונות במשקעים בין שתי השנים, דומות המגמות של השפעת כתמי היער השונים וההצללה המלאכותית על שיעורי ההתבססות. תוצאות אלו עומדות בניגוד להשערת הניסוי ול"השערת ההקלה" הטוענת, כי ככל שהעקה חמורה יותר ההקלה תהיה משמעותית יותר [8].
Zavala טוען [35], כי הדינמיקה של היער המעורב באזורים ים-תיכוניים אינה מוכתבת על ידי זמינות האור אלא על ידי זמינות המים. זאת, בניגוד לדינמיקת היער המעורב הממוזג. מחקר זה מלמד, כי שני הגורמים חשובים לדינמיקת האוכלוסייה של האלון המצוי, אבל תרומתם היחסית עשויה להשתנות במשך מחזור החיים של המעוצה המתבסס ואולי אף להשפיע על דינמיקת ההיווצרות של היער המעורב.
תודות
המחקר מומן על ידי הקרן הקימת לישראל ואוניברסיטת בן-גוריון. בית הספר ללימודי מדבר, המכונים לחקר המדבר, אוניברסיטת בן-גוריון, השתתף בתמיכה בשני בן-יאיר בביצוע עבודת המחקר.תודה מיוחדת לארנון ברנד ולעוזרי המחקר הרבים אשר השתתפו בהקמת מערך הניסוי ובתצפיות. תודות לאלי בן-שיטרית, לעדי נעלי ולרבים מאנשי קק"ל על תמיכתם במחקר זה.
מקורות
- אשד י. 1991. ריבוי אלונים מקומיים מבלוטים ויחורים. עבודת מוסמך, האוניברסיטה העברית.
- גולדרינג י. 1977. התחדשות הצומח הטבעי ביערות אורן נטועים. עבודת מוסמך, האוניברסיטה העברית.
- חריף י. 1974. התפתחות מרכיבי הגריגה והחורש הראשיים בשנתם הראשונה וחשיבותם בקביעת הטורים הסוקצסיוניים בהרי יהודה. חיבור לשם קבלת תואר דוקטור לפילוסופיה, האוניברסיטה העברית.
- טפר א. 1997. הגורמים המשפיעים על עיצוב צורת הגידול של האלון המצוי. עבודת מוסמך, האוניברסיטה העברית.
- רביקוביץ ש. 1981. קרקעות ישראל: התהוותן, טבען ותכונותיהן. הוצאת הקיבוץ המאוחד.
- Alon G and Kadmon R. 1996. Effect of successional stages on the establishment of Quercus calliprinos in an East Mediterranean maquis. Israel Journal of Plant Sciences 44: 335-345.
- Bell S and Apostol D. 2007. Designing Sustainable Forest Landscapes. Taylor and Francis.
- Bertness MD and Callaway R. 1994. Positive interactions in communities. Trends Ecol Evol 9: 191-193.
- Brokaw N and Busing RT. 2000. Niche versus chance and tree diversity in forest gaps. Trends Ecol Evol 15: 183-188.
- Callaway RM and Davis FW. 1998. Recruitment of Quercus agrifolia in central California: The importance of shrub-dominated patches. J Veg Sci 9: 647-656.
- Clark JS, Beckage B, Camill P, Cleveland B, HilleRisLambers J, Lighter J, McLachlan J, Mohan J, and Wyckoff P. 1999. Interpreting recruitment limitation in forests. American Journal of Botany 86: 1-16.
- Clinton BD. 2003. Light, temperature, and soil moisture responses to elevation, evergreen understory, and small, canopy gaps in the southern Appalachians. Forest Ecology and Management 186: 243-255.
- Endler JA. 1993. The Color of Light in Forests and Its Implications. Ecological Monographs 63: 1-27.
- Flores J and Jurado E. 2003. Are nurse-protege interactions more common among plants from arid environments? J Veg Sci 14: 911-916.
- Glenn-Lewin DC, Peet RK, and Veblen TT. 1992. Plant Succession :Theory and Prediction. Chapman and Hall.
- Grime JP. 1966. Shade avoidance and shade tolerance in flowering plants. Eds. Bainbridge R, Evans GC, and Rackham O., Blackwell.
- Holmgren M, Scheffer M, and Huston MA. 1997. The interplay of facilitation and competition in plant communities. Ecology 78: 1966-1975.
- Maestre FT, Cortina J, Bautista S, and Bellot J. 2003. Does Pinus halepensis facilitate the establishment of shrubs in Mediterranean semi-arid afforestations? Forest Ecology and Management 176: 147-160.
- Miriti MN. 2006. Ontogenetic shift from facilitation to competition in a desert shrub. J of Ecol 94: 973-979.
- Nathan R and Muller-Landau HC. 2000. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol Evol 15: 278-285.
- Niinemets U and Valladares F. 2006. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecological Monographs 76: 521-547.
- Osem Y, Ginsberg P, Tauber I, Atzmon N, and Perevolotsky A. 2008. Sustainable management of Mediterranean planted coniferous forests: An Israeli definition. Journal of Forestry 106: 38-46.
- Prider JN and Facelli JM. 2004. Interactive effects of drought and shade on three arid zone chenopod shrubs with contrasting distributions in relation to tree canopies. Functional Ecology 18: 67-76.
- Reisman-Berman O. 2007. Age-related change in canopy traits shifts conspecific facilitation to interference in a semi-arid shrubland. Ecography 30: 459-470.
- Ricklefs RE. 1977. Environmental heterogeneity and plant species-diversity – hypothesis. Am Nat 111: 376-381.
- Rodriguez-Calcerrada J, Pardos JA, Gil L, and Aranda I. 2008. Ability to avoid water stress in seedlings of two oak species is lower in a dense forest understory than in a medium canopy gap. Forest Ecology and Management 255: 421-430.
- Sanchez-Gomez D, Valladares F, and Zavala MA. 2006. Performance of seedlings of Mediterranean woody species under experimental gradients of irradiance and water availability: trade-offs and evidence for niche differentiation. New Phytologist 170: 795-805.
- Sanchez-Gomez D, Zavala MA, and Valladares F. 2006. Seedling survival responses to irradiance are differentially influenced by low-water availability in four tree species of the Iberian cool temperate-Mediterranean ecotone. Acta Oecol Int J Ecol 30: 322-332.
- Smith T and Huston M. 1989. A theory of the spatial and temporal dynamics of plant communities. Vegetatio 83: 49-69.
- Thomas P and Packham J. 2007. Ecology of Woodlands and Forests: Description, Dynamics and Diversity. Cambridge University Press.
- Valladares F and Pearcy RW. 1997. Interactions between water stress, sun-shade acclimation, heat tolerance and photoinhibition in the sclerophyll Heteromeles arbutifolia. Plant Cell Environ 20: 25-36.
- Valladares F and Pearcy RW. 2002. Drought can be more critical in the shade than in the sun: a field study of carbon gain and photo-inhibition in a Californian shrub during a dry El Nino year. Plant Cell Environ 25: 749-759.
- Valladares F and Niinemets U. 2008. Shade tolerance, a key plant feature of complex nature and consequences. Annual Review of Ecology Evolution and Systematics 39: 237-257.
- Verheyen K and Hermy M. 2001. The relative importance of dispersal limitation of vascular plants in secondary forest succession in Muizen Forest, Belgium. J Ecol 89: 829-840.
- Zavala MA, Espelta JM, and Retana J. 2000. Constraints and trade-offs in Mediterranean plant communities: The case of Holm oak-Aleppo pine forests. Bot Rev 66: 119-149.




 מהדורה מודפסת
מהדורה מודפסת








