
סקירות
התנוונות ותמותה מוקדמת של אורן ירושלים בישראל בעקבות יבוא זרעים ממערכות אקולוגיות זרות

המאמר מסכם את ההיסטוריה של יבוא גנוטיפים זרים של אורן ירושלים במסגרת מפעל הייעור, את המחקר הגנטי בנושא ואת ההשלכות של הגנוטיפים הזרים על היער | צילום: דודי שמחון
אורן ירושלים הוא מין מועדף למטרות ייעור ושיקום שטחים מופָרים, לא רק בישראל אלא בכל רחבי אגן הים התיכון ומעבר לו. הסיבות לכך מרובות, ובהן הטיפול הפשוט בו במשתלה, פשטות נטיעתו, היקלטותו הטובה, גדילתו המהירה, עמידותו ליובש יותר מאשר אורנים ים תיכוניים אחרים, איכות העצה שלו, שאינה נופלת בטיבה מהאיכות של אורנים אחרים, והיותו מין חלוץ בשטחים מופָרים. בעקבות התנוונות ותמותה של אורן ירושלים ביערות נטועים מבלי שנמצאה לכך סיבה אקולוגית ברורה וחד-משמעית, החל מחקר גנטי מתקדם במין זה על פני מרחב תפוצתו באגן הים התיכון.
תוצאות המחקר הוכיחו שאף על פי שאורן ירושלים גדֵל באופן טבעי באזורנו כבר לפחות שניים וחצי מיליון שנה, כמעט שלא נאספו זרעים ביערות הטבעיים בארץ למטרות ריבוי בתקופת המנדט הבריטי וגם לא אחריו. הזרעים יובאו, מפאת הנוחיות שבדבר, בעזרת חברות זרעים מארצות ים תיכוניות אחרות, שהתנאים האקולוגיים בהן שונים משל ישראל, ולכן גם ההרכב הגנטי של אוכלוסיות האורן בהן שונה מזה של האוכלוסיות הטבעיות בישראל. לאחר מכן נאספו זרעים ביערות הנטועים הראשונים. נוהג זה של יבוא זרעים הוא בניגוד גמור לכלל היערני שלפיו אין להפיץ זרעים אל תחום אקולוגי אחר. כנראה שחוסר התאמה בין המבנה הגנטי של האקוטיפים שיובאו לבין התנאים האקולוגיים בשטחים שיוערו כאן, הוא אחד מהגורמים שגרמו להתנוונות ולתמותה מוקדמת יחסית של רבים מיערות אורן ירושלים הנטועים בארץ. גורם נוסף להתנוונות הוא טיפול שאינו מקצועי דיו ביער, כתוצאה ממחסור בידע מקצועי ובכוח אדם מקצועי בקק"ל. כללים שגובשו בשנים האחרונות באגף הייעור של קק"ל תורמים לטיפול מושכל ביערות הצעירים.
נוסף על כך, כתוצאה מההאבקה גם על-ידי אבקה מעצים נטועים, שהגנום בהם כולו או חלקו הוא מערב ים תיכוני, נוצר זיהום של המאגר הגנטי של אורן ירושלים הטבעי במדינה, וכך המגוון הגנטי של המין הזה הולך וקטֵן.
על קצה המזלג
- החל בתקופת המנדט הבריטי יוערו שטחים נרחבים באמצעות שתילת אורן ירושלים.
- אף על פי שאורן ירושלים גדל באזורנו לפחות שניים וחצי מיליון שנה, יובאו זרעיו מארצות ים תיכוניות אחרות למטרות ריבוי, וזאת מטעמי נוחות.
- השוני בתנאים האקולוגיים בין ארצות אלה לישראל יצר חוסר התאמה בין המבנה הגנטי של האקוטיפים שיובאו ובין התנאים האקולוגיים בשטחים שיוערו בארץ. הדבר תרם להתנוונות עצים ולתמותה מוקדמת שלהם.
- זיהום המאגר הגנטי הטבעי של אורן ירושלים וטיפול שלא היה מקצועי דיו ביערות בעבר, הובילו לכך שהמגוון הגנטי של המין הולך וקטֵן.
- איסוף סלקטיבי של זרעים ממקורות נבחרים, כמו גם טיפול מושכל ביער הצעיר, צפויים לשפר את עמידות האורנים לתנאי הארץ.
המערכת
הלכה למעשה
האורן כחלק מהפלורה של ישראל
אורן ירושלים (Pinus halepensis Mill) או בשמו האנגלי Aleppo pine הוא המין היחידי ממשפחת האורנים שגדל באזור שהוא כיום ישראל וירדן, לפחות מתקופת הפליסטוקן (שניים וחצי מיליון שנה) [24]. כלומר, מין זה הוא חלק מובנה מהצומח הטבעי בישראל [53]. מההבדלים בכמות גרגירי האבקה של אורן ירושלים שנמצאו בשכבות קרקע שהוצאו מעומקים שונים בקידוחים יבשתיים וימיים, אפשר להסיק על תנודות גדולות יחסית בנוכחותו המרחבית בתקופות גֵאולוגיות והיסטוריות שונות [52].
בשנת 1880 פורסמו תוצאות הסקר והמיפוי שהתבססו על המדידות של משלחת החברה הבריטית לחקירת ארץ ישראל [15]. במפות אלה קיים סימון ברור להימצאותם של עצי מחט (אורן ירושלים) בהיקף קטן במספר מצומצם של מקומות: ירכא, הכרמל, אזור אום-צפא בשומרון ובית מאיר (שמורת "המסרק"). במפות מאוחרות יותר (הסקרים של ארץ ישראל שפורסמו בשנים 1937-1934) גדל מספר המקומות שסומן בהם קיומם של עצי אורן טבעיים [3].
מקורות זרעים אקזוטיים של אורן ירושלים ביערות הנטועים
התנוונות ותמותה של עצים רבים ביער שער הגיא בשנת 1972/3 עוררה את התעניינות הציבור, שדרש הסברים לתופעה באמצעות העיתונות. דרישה זו יצרה גל של מחקרים בתחומים רבים ושונים של מדעי הצמח, מדעי הקרקע וגאולוגיה, השפעת זיהום אוויר, השפעת ממשק היער, מזיקים ומחלות ועוד, שניסו להסביר את התופעה [1, 4]. במסגרת זו נעשה גם מחקר על התפתחות העצים שניטעו ביער שער הגיא בין השנים 1936-1926 על-ידי אגף הייעור המנדטורי, תחת השפעת תנאים אקולוגיים מקומיים. במהלך הסקר אובחנו ביער לפחות שלושה מופעים (פנוטיפים – phenotypes) של אורן ירושלים, השונים מאוד זה מזה בצורת הנוף, בכמות האצטרובלים, בצורת הקליפה ועוד. עצים ממופע מסוים לא נפגעו, בעוד שעצים בעלי מופע אחר נפגעו מאוד [2]. בדיקות מורפולוגיות של קליפת העצים והמחטים ושל הרכב השרף בעצה (כסמן גנטי) בכל אחד משני המופעים (מופע פגוע ומופע שאינו פגוע) ובעצים הגדלים במסרק שנחשבו טבעיים מקומיים, הוכיחו כי כל קבוצת עצים שונה מאוד מחברתה [38]. התוצאות ממחקר זה האיצו סקירה בארכיון של אגף הייעור המנדטורי השמור בגִנזך המדינה כדי להבין את מה שאירע בעבר. נמצאו התכתבויות, חשבוניות והוראות תשלום לחברות הזרעים וויל-מורן מצרפת וגרינוולד מאוסטריה עבור אספקת זרעי אורן ירושלים בכמויות גדולות. המסקנה העולה מממצא זה היא שרק מיעוט מהזרעים שהיו דרושים לגידול השתילים עבור נטיעת היערות נאספו בחלקות היער הטבעיות (עומדים) של אורן ירושלים בארץ, כלומר מאקוטיפים (אוכלוסיות מקומיות בתוך מין ביולוגי בעלות מערך התאמות גנטי מסוים לתנאי סביבה ייחודיים – ecotypes) המותאמים לתנאים האקולוגיים המקומיים בישראל. רוב הזרעים הגיעו מעבר לים – מסביבות אקולוגיות ששונות לחלוטין מאלה של ישראל. ליבוא זה של זרעים מאקוטיפים שהם זרים לאקולוגיה המקומית, הייתה וקיימת עדיין השפעה רבה על היערות שניטעו בעבר, וגם על ההרכב הגנטי של האוכלוסיות הטבעיות והייחודיות של אורן ירושלים טבעי בישראל [49]. אם משלוחי הזרעים שהגיעו לארץ הכילו מידע גאוגרפי ואקולוגי על מקורם הרי שהוא לא נשמר, ולכן עלתה השאלה – מהיכן הזרעים? מדובר בשאלה משמעותית ביותר בתחום היערנות, היות שהעצים הניטעים חייבים לשרוד ולהתפתח במשך עשרות שנים רבות מאוד ללא השקיה, דישון ופעולות נגד מזיקים ומחלות, כי הרי יערנות אינה חקלאות או גננות.
המגוון הגנטי באורן ירושלים מעבריו השונים של הים התיכון
כדי לנסות ולענות על השאלה "מהיכן הזרעים" נעשו בדיקות גנטיות שכללו: בדיקות של הרכב השרף, שהופק מהעצה ומהעלווה של אוכלוסיות טבעיות ונטועות בארץ [6, 42, 43]; בדיקות של המגוון הגנטי על-ידי שימוש באיזוזימים (חלבונים תוצרי גנים – isozymes). בדיקות אלה נעשו בזרעים שמקורם באוכלוסיות טבעיות רבות מסביב לאגן הים התיכון [35, 44], מיערות טבעיים בארץ ומיערות שנשתלו בזמן המנדט הבריטי ולאחר קום המדינה [23].
הוכחה לכך שאכן נשתלו בארץ עצים מאקוטיפים ששונים מאלה של האורנים הטבעיים בישראל נובעת מהבדלים כמותיים בחומרים המרכיבים את השרף [43] ומהבדלים בנוכחות ובתדירות של אללים בגנים שונים בעצים שגדלים ביערות נטועים בארץ; אללים אלה ייחודיים לאזורים גאוגרפיים אחרים ושונים סביב אגן הים התיכון, ואינם בנמצא באוכלוסיות טבעיות בארץ [23, 44, 49].
התוצאות שמובאות באיור 1א מצביעות על השינוי (הירידה) במגוון הגנטי שמבוטא כהטרוזיגוטיות הממוצעת באקוטיפ, כלומר באוכלוסייה שהייתה המקור לזרעים לאורכו של אגן הים התיכון, מאזור הבלקנים במזרח ועד לספרד ומרוקו במערב (על פי קווי אורך גאוגרפיים). בתוצאות אלה תומכת שורה ארוכה של בדיקות גנטיות אחרות [7, 23, 25, 30, 44]. איור 1ב מראה כי קיימת נטייה לעלייה במגוון הגנטי באוכלוסיות הטבעיות של אורן ירושלים מצפון כלפי דרום באזור מזרח אגן הים התיכון (ישראל וירדן), בדומה לעלייה במידת הצחיחות מצפון לדרום ולהשפעתה על המגוון הגנטי במיני צמחים ובעלי חיים [36, 37]. נטייה זו אינה מובהקת מבחינה סטטיסטית. הירידה או העלייה במגוון הגנטי (מבוטא כהטרוזיגוטיות) קשורה למספר הגנים הפולימורפיים בכל עץ ולתדירות של האללים השונים בגנים אלה באוכלוסייה.
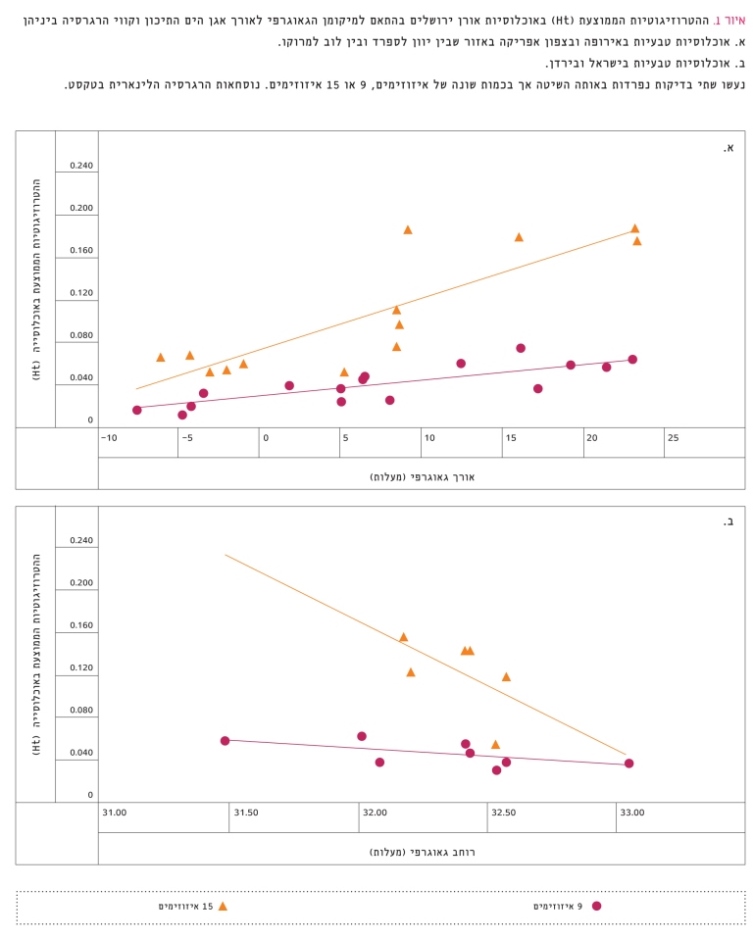
איור 1
ההטרוזיגוטיות הממוצעת (Ht) באוכלוסיות אורן ירושלים בהתאם למיקומן הגאוגרפי לאורך אגן הים התיכון וקווי הרגרסיה ביניהן
א. אוכלוסיות טבעיות באירופה ובצפון אפריקה באזור שבין יוון לספרד ובין לוב למרוקו.
ב. אוכלוסיות טבעיות בישראל ובירדן.
נעשו שתי בדיקות נפרדות באותה השיטה אך בכמות שונה של איזוזימים, 9 או 15 איזוזימים. נוסחאות הרגרסיה הלינארית בטקסט.
בעשור האחרון נוספו תוצאות ממחקרים רבים שנעשו בדי-אן-איי שבגרעין, בכלורופלסטים ובמיטוכונדריות. התוצאות מחזקות את ההבנה בדבר הרחבת אזור התפוצה של אורן ירושלים בתקופת הפליסטוקן וההולוקן ממקום היווצרותו המשוער [44] שבצפון-מזרח אגן הים התיכון (כנראה האזור שבין הים השחור והים הכספי), לדרום מערב (ספרד, מרוקו). השערה זו מבוססת על הקִרבה הגנטית בין אורן ירושלים לאורן ברוטיה (Pinus brutia Ten) על ארבעת תתי-המין שבו. יש הטוענים שלתנאי האקלים בשיא תקופת הקרח האחרונה, שהיו שונים מאוד לאורכו של אגן הים התיכון, הייתה השפעה רבה על הפחיתה במגוון הגנטי לאורכו של האגן. נוסף על כך השפיע גם שוני בתנאים הגאולוגיים והטופוגרפיים, שיצרו את מה שמכונה "צוואר בקבוק" – גורם אקלימי, טופוגרפי או אקולוגי אחר המצמצם מאוד את גודל האוכלוסיות, ומכאן גם את המגוון הגנטי שלהן. דוגמה ל"צוואר בקבוק" היא למשל המגוון הנמוך באוכלוסיות במרוקו לעומת המגוון באוכלוסיות שגדלות בשאר המדינות בצפון אפריקה [8, 18, 19, 20, 21, 22, 40].
אקוטיפים של אורן ירושלים ועמידות ליובש
העמידות של צמחים ים תיכוניים למיעוט מים בקרקע בתקופת הקיץ, שעשויה להתארך למעל ל-250 ימים ללא גשמים במזרח אגן הים התיכון, העסיקה חוקרים רבים. נמצא כי מבין מיני האורנים הים תיכוניים, אורן ירושלים הוא העמיד ביותר ליובש כתוצאה מתכונותיו האקופיזיולוגיות [33, 39, 46]. מסוף שנות ה-70 של המאה ה-20 החלו לחקור הבדלים לא רק ברמת המין אלא גם ברמת האוכלוסיות בתוך המין, שנבדלות בתנאי בית הגידול שלהן. הבדלים בין אקוטיפים נמצאו בקצב הנביטה והתפתחות הנבטים בהשפעת הפוטנציאל האוסמוטי של מצע ההנבטה והגידול, בהשפעת עוצמת האור ובהשפעת הטמפרטורה [10, 11, 12, 13]. לאורך השנה ההידרולוגית נמצאו הבדלים יומיים בין אקוטיפים שגודלו בתנאים אחידים, בפוטנציאל המים בעלווה לפני הזריחה ובצהרי היום, ברמת הגירעון לרוויה של המחטים, ברמת הדיות והפוטוסינתזה. כלומר, הבדלים ביחס שבין הדיות לקיבוע פחמן דו-חמצני, ומכאן הבדלים בעמידות ליובש, בקצב הצמיחה ובתכונות פיזיולוגיות נוספות וגם בעמידות למזיקים ולמחלות [5, 7, 25, 26, 41, 47, 48, 50].
נוסף על כך, מחקרים על תגובת שתילי אורן ירושלים לתנאי יובש הראו כי השתילים שמוצאם מאזורים שקיימים בהם תנאי יובש, עמידים יותר לעקת מים במידה מובהקת. בהשוואה לשתילים שמוצאם מאזורים לחים יותר, לשתילים אלה יש מוליכות מים גבוהה יותר בין הקרקע למחטים, מוליכות מים גבוהה יותר במחטים, דיות רבה יותר, התאמה אוסמוטית טובה יותר וגם רמה נמוכה יותר של חדירת אוויר לצינורות ההובלה [13, 50]. נוסף על כך, עצי אורן ירושלים בני אותו גיל, שמוצאם מאוכלוסיות שונות מסביב לאגן הים התיכון, שגודלו בחלקת ניסוי אחידה באזור צחיח (יער יתיר), נבדלו באופן מובהק ברמת פוטנציאל המים בעצה של ענפונים ושל מחטים שנמדדה לפני הזריחה [48]. בדיקות אנטומיות בעצה של עצים מאקוטיפים שונים, שגודלו בתנאים אחידים במשתלה או בחלקת ניסוי, הראו שוני במבנה האנטומי של העצה בהתאם לתנאים האקולוגיים באוכלוסיות המוצא. הצמחים נבדלו באפשרויות למעבר מים (אורך צינורות ההובלה וקוטרם בשורשים ובגזע, גודל הגומָצים [הפתחים שבמקום החיבור של צינורות ההובלה]), ומכאן גם בעמידות ליובש [17]. בדיקות של התפתחות שתילי אורן ירושלים שמוצא הזרעים שלהם הוא בארבע אוכלוסיות בספרד, שנבדלות בתנאים האקולוגיים שהתפתחו בהם, ושגודלו במשתלה בתנאים אחידים, הראו כי קיימים הבדלים מובהקים בין האוכלוסיות בתגובת השתילים למשטר ההשקיה. ההבדלים התבטאו במשקל היבש של השורשים, של הגזע, של המחטים ושל כל השתיל, בגובה השתיל, בקוטר הגזע ובצמיחה לגובה במשך תקופת ההימנעות מהשקיה [14]. עקת מים באביב ובתחילת הקיץ עשויה לגרום שינויים אנטומיים במבנה הטבעת השנתית שמעכב מעבר מים [16]. לאחרונה הוכח גם כי עקה זו עשויה להשפיע מאוד על מידת הקליטה של שתילים בנטיעות ועל התחדשות טבעית של יערות.
אקוטיפים של אורן ירושלים והתאמתם לתנאי הארץ
כדי להשוות בין מקורות זרעים שונים של אורן ירושלים במידת העמידות והצמיחה של השתילים בתנאים אקולוגיים שונים בארץ, נשתלו בשנת 1976 שלוש חלקות ניסוי: ברמת הנדיב, ביער הקדושים וביער נחשון [51]. איור 2 מתאר את היחס שבין אחוז ההישרדות של העצים במסגרת האקוטיפים השונים 10 שנים לאחר הנטיעה לבין ההטרוזיגוטיות הממוצעת של אקוטיפים אלה שנטועים בכל אחת משלוש חלקות הניסוי. הרגרסיה הלינֵארית שבין רמת ההטרוזיגוטיות ואחוז ההישרדות בכל החלקות היא מובהקת:
- ברמת הנדיב: r2=0.390, p<0.030, n=12
- ביער נחשון: r2=0.410, p<0.026, n=12
- ביער הקדושים: r2=0.460, p<0.0015, n=12
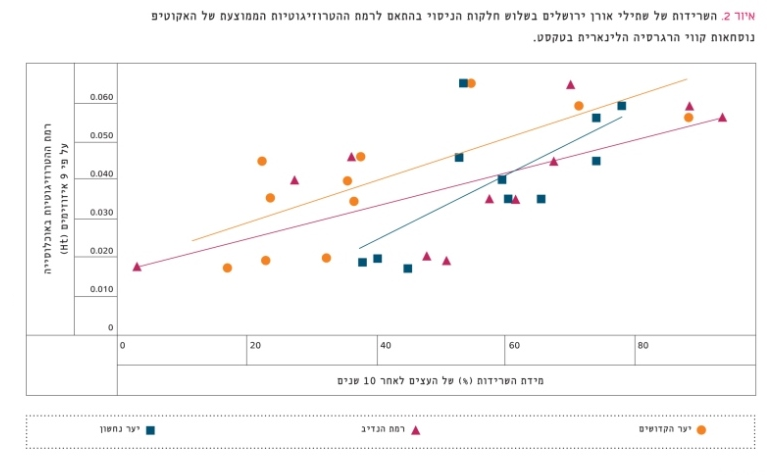
איור 2
השרידות של שתילי אורן ירושלים בשלוש חלקות הניסוי בהתאם לרמת ההטרוזיגוטיות הממוצעת של האקוטיפ
נוסחאות קווי הרגרסיה הלינארית בטקסט
איור 3 מתאר את היחס שבין מידת העמידות למָצוּקוֹקוּס (Matsucoccus josephi) [34], שהיא כנימה מזיקה של אורן ירושלים, לבין ההטרוזיגוטיות הממוצעת באוכלוסיות השונות הנטועות בשתי חלקות ניסוי. הרגרסיה הלינארית בין שני המשתנים הללו בחלקת רמת הנדיב היא מובהקת (r2=0.786, p<0.0006, n=10), וכך גם בחלקת נחשון (r2=0.445, p<0.035, n=10). כלומר, מאיור 2 ואיור 3 נראה בבירור כי ככל שגדלה ההטרוזיגוטיות הממוצעת באוכלוסייה כן עולות השרידות והעמידות למצוקוקוס, ממצא התואם את הניסיון והידע היערניים [27, 28, 32]. אפשר רק להצטער שבחלקות הניסוי הללו לא נשתלו גם שתילים שמוצאם מהיערות הטבעיים בארץ. במחקר נוסף בארץ על הבדלים בין אוכלוסיות נמצא כי הצמיחה והתגובה לתנאי יובש קשורות לרמת ההטרוזיגוטיות [45].
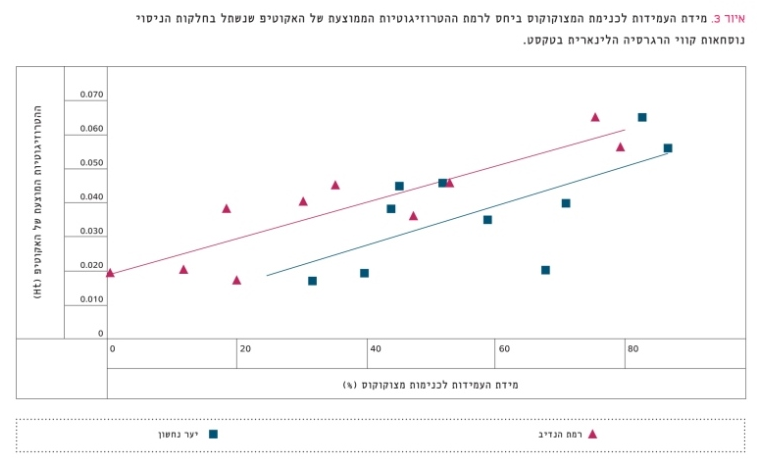
איור 3
מידת העמידות לכנימת המצוקוקוס ביחס לרמת ההטרוזיגוטיות הממוצעת של האקוטיפ שנשתל בחלקות הניסוי
נוסחאות קווי הרגרסיה הלינארית בטקסט
סיכום ומשמעויות
תוצאות ממחקרים פָּלינולוגיים [24, 52] הוכיחו שאורן ירושלים גדל באופן טבעי בישראל כבר לפחות שניים וחצי מיליון שנה, דבר שהביא להתאמה גנטית לאקולוגיה כאן. כאשר אין פוגעים בו ובית הגידול תואם את דרישותיו האקולוגיות, הוא עשוי להתקיים ולחיות עד גיל שבין מאה למאתיים שנים (אוכלוסיית המסרק, עצים בכרמל, עצים בעיר העתיקה בירושלים ובחברון). לצערנו כמעט שלא נאספו זרעים ביערות הטבעיים בארץ בתקופת המנדט הבריטי ולאחריו. הזרעים יובאו, מפאת הנוחיות, בעזרת חברות זרעים מאזורים גאוגרפיים ומתנאים אקולוגיים שונים משל ישראל; לכן ההרכב הגנטי – המושפע מהתנאים האקולוגיים שבמקום מוצא הזרעים – של רבים מיערות אורן ירושלים שניטעו בישראל, שונה מזה של האוכלוסיות הטבעיות כאן [44, 47]. נוהג זה של יבוא זרעים הוא בניגוד גמור לכלל היערני שלפיו אין להפיץ זרעים מתחום אקולוגי אחד למשנהו ללא בדיקה מעמיקה, שלא התקיימה בעבר. ביערנות קיים המושג "אזור תפוצת זרעים" (seed dispersal zone) כלומר, האזור שמותר להעביר אליו זרעים ממטעי האֵם לזרעים ו/או למחלקות יער נבחרות, היות שהתנאים האקולוגיים במקומות הללו דומים מאוד. כנראה שחוסר ההתאמה בין המבנה הגנטי של האקוטיפים שיובאו לישראל לבין התנאים האקולוגיים בשטחים שיוערו, הוא אחד מהגורמים להתנוונות ולתמותה מוקדמת יחסית של רבים מיערות האורן הנטועים בארץ.
רק מקור אחד לזרעים של אורן ירושלים מחו"ל הוכיח את עצמו מאז ימי המנדט הבריטי ועד עתה בצמיחתו ובעמידותו ליובש ולמזיקים. מוצאו מחצי האי הבלקני (ולכן הוא מכונה אורן ירושלים יווני), והוא שונה במבנה ובמגוון הגנטי מכל שאר האוכלוסיות במרחבי אגן הים התיכון [7, 31, 34, 44, 47, 48]. נוסף על כך, אין להמעיט בהשפעה השלילית של חוסר טיפול (דילול) במועד ובעוצמה הנדרשים בהתאם לגיל היער, למידת התפתחותו ולתנאי בית הגידול, דבר שגרם לתחרות עזה שהובילה להתנוונות עצים. תופעה זו נבעה מחוסר הבנה של הדינמיקה של התפתחות עצים ויער ושל השפעת התחרות על דינמיקה זו, ומהמחסור החמור בכוח אדם מקצועי. עתה מודעים באגף הייעור של קק"ל לצורך באיסוף זרעים ממקורות נבחרים כמו גם לצורך בטיפול ביער הצעיר בצורה מושכלת, ומנסים לבצע מטלה זו על פי הכללים שגובשו בשנים האחרונות באגף.
ייצור זרעים מושבחים עבור נטיעת יערות חדשים
המבנה הגנטי נבדק בשיטה של הרצת איזוזימים במצע עמילן בשדה חשמלי. בפעם הראשונה נבדקו 9 איזוזימים, ובפעם השנייה 15 [30, 44]. בהתאם לבדיקות נמצא כי ההטרוזיגוטיות הממוצעת של אורן ירושלים טבעי מזרח ים תיכוני (בארץ ובירדן) ובחצי האי הבלקני בבדיקה הראשונה הייתה 0.046 ו-0.064, ובבדיקה השנייה הייתה 0.123 ו-0.183, בהתאמה. על פי רמת ההטרוזיגוטיות ניתן להניח בסבירות גבוהה שצאצאים מהאוכלוסיות הטבעיות של אורן ירושלים במזרח אגן הים התיכון יהיו בעלי שרידות גבוהה ועמידות גבוהה למצוקוקוס.
ידע מקצועי בתחום היערנות קושר בין מידת העמידות לתנאי סביבה קשים ומידת העמידות למזיקים ולמחלות לבין רמת ההטרוזיגוטיות של הפרט, כלומר, לכמות הגנים הפולימורפיים שקיימים בו [9, 27, 28, 32]. באורן ירושלים מחצי האי הבלקני ובאורן ירושלים טבעי בישראל קיימת הטרוזיגוטיות שגבוהה משאר האוכלוסיות הים תיכוניות [29, 31, 44], אך המבנה הגנטי בשתי הקבוצות שונה (אותם גנים הם מונומורפיים או פולימורפיים באחת משתי הקבוצות). ידע זה היה הבסיס להחלטה משותפת לאגף הייעור ולהנהלת ענף היער בהובלתו של פרופ' דניאל זהרי (גנטיקאי) להקים חמישה "מטעי-אֵם" לזרעים באזורים אקולוגיים שונים במדינה, כדי שהזרעים שיתקבלו ישמשו לנטיעת יערות חדשים ולשיקום יערות מתנוונים שאינם מתחדשים מזריעה עצמית עקב תנאים אקולוגיים צחיחים. במטעים אלה נשתלה תערובת של אורן ירושלים טבעי מקומי ומחצי האי הבלקני, כדי שהפריה הדדית תתבטא בהעלאת ההטרוזגוטיות.
שיקום היערות המבוגרים המתנוונים ללא נטיעה מחדש
שיקום יערות בשלבי התנוונות כתוצאה מחוסר התאמה גנטית לתנאי בית הגידול אפשרי גם בעזרת חידוש טבעי, שבמסגרתו מתקיימת ברֵרה טבעית בין הנבטים והזריעים בהתאם לתנאים האקולוגיים בכל נישה. בסופו של דבר, לאחר מחזורי חידוש טבעי אחדים, תשאיר הברֵרה הטבעית גנומים המותאמים טוב יותר לתנאי בית הגידול.
הנטיעה בעבר של אקוטיפים שאינם מזרח ים תיכוניים מסכנת את המבנה הגנטי העתידי של העומדים הטבעיים של אורן ירושלים שעוד נותרו במדינה, כתוצאה מפיזור אבקה למרחקים [49]. שימור הגנום המזרח ים תיכוני על מגוון האקוטיפים שבו, ששונים במבנה הגנטי שלהם משאר האוכלוסיות סביב אגן הים התיכון, יכול להיעשות במספר שיטות המשלימות אחת את השנייה: א. שימור במקום (in situ) בעזרת ממשק יער מתוכנן לכך, שיפריד עומדים טבעיים בעזרת אזורי חיץ, דבר שעשוי להפחית את רמת ההפריה החיצונית בעומד; ב. שימור בעזרת בנק הגנים (ex situ); ג. שימור בעזרת ריבוי וגטטיבי ונטיעה באזור נקי יחסית מיערות נטועים.
- החל בתקופת המנדט הבריטי יוערו שטחים נרחבים באמצעות שתילת אורן ירושלים.
- אף על פי שאורן ירושלים גדל באזורנו לפחות שניים וחצי מיליון שנה, יובאו זרעיו מארצות ים תיכוניות אחרות למטרות ריבוי, וזאת מטעמי נוחות.
- השוני בתנאים האקולוגיים בין ארצות אלה לישראל יצר חוסר התאמה בין המבנה הגנטי של האקוטיפים שיובאו ובין התנאים האקולוגיים בשטחים שיוערו בארץ. הדבר תרם להתנוונות עצים ולתמותה מוקדמת שלהם.
- זיהום המאגר הגנטי הטבעי של אורן ירושלים וטיפול שלא היה מקצועי דיו ביערות בעבר, הובילו לכך שהמגוון הגנטי של המין הולך וקטֵן.
- איסוף סלקטיבי של זרעים ממקורות נבחרים, כמו גם טיפול מושכל ביער הצעיר, צפויים לשפר את עמידות האורנים לתנאי הארץ.
המערכת
מקורות
- שילר ג. 1974. תולדותיו של יער שער-הגיא. ליערן 24(3–4): 48–52.
- שילר ג. 1977. יחסי גומלין שבין התפתחות אורן ירושלים לבין תנאי הסביבה. ליערן 27(1–4): 13–23.
- שילר ג. 1985. אורן ירושלים טבעי: תפוצה וקשרים גנטיים. רתם 18: 69–78.
- שילר ג. 2003. אוזון ואורן ירושלים. יער 4: 60–66.
- Atzmon N, Moshe Y, and Schiller G. 2004. Ecophysiological response to sever drought in Pinus halepensis trees of two provenances. Plant Ecology 171: 15–22.
- Baradat Ph, Michelozzi R, Tognetti R, et al. 1995. Geographical variation in the terpene composition of Pinus halepensis In: Baradat Ph, Adams WT, and Mueller-Starck G (Eds). Population genetics and genetic conservation of forest trees. Amsterdam: SPB Academic Publishing.
- Bariteau M, Huc R, and Vendramin GG (Coordinators). 2000. Mediterranean Pinus and Cedrus (MPC): Adaptation and selection of Mediterranean Pinus and Cedrus for sustainable afforestation of marginal lands. Commission of the European Communities, Directorat General for Agriculture, DGVI FII 3, Contract FAIR CT95-0097.
- Bucci G, Anzidei M, Madaghiele A, and Vendramin GG. 1998. Detection of haplotypic variation and natural hybridization in halepensis-complex pine species using chloroplast simple sequence repeat (SSR) markers. Molecular Ecology 7: 1633–1643.
- Bush R and Smouse PE. 1987. The fitness consequences of multiple-locus heterozygosity: The relationship between heterozygosity and growth rate in Pitch pine (Pinus rigida). Evolution 41(4): 787–798.
- Calamassi R, Falusi M, and Tocci A. 1980. Variazione geografica e resistenza a stress idrici in semi di Pinus halepensis, Pinus brutia Ten. e Pinus eldarica Medw. Annali Instituto Sperimentale per la selvicoltura di Arezzo 11: 194–230.
- Calamassi R. 1982. Effetti della luce e della temperature sulla graminazione dei semi in provenienze di Pinus halepensis e Pinus brutia Ten. L' Italia Forestale e Montana 37: 174–187.
- Calamassi R, Paoletti E, and Starati S. 2001a. Frost hardening and resistance in three Aleppo pine (Pinus halepensis) provenances. Israel Journal of Plant Sciences 49: 179–186.
- Calamassi R, Rocca GD, Falusi M, et al. 2001b. Resistance to water stress in seedlings of eight European provenances of Pinus halepensis. Annals of Forest Science 58: 663–672.
- Chambel MR, Climent J, and Alia R. 2007. Divergence among species and populations of Mediterranean pines in biomass allocation of seedlings grown under two watering regimes. Annals of Forest Science 64: 87–97.
- Conder FR, and Kitchener HH. 1880. Map of Western Palestine, in 24 Sheets, from the Surveys Conducted for the Committee of Palestine Exploration Fund. London: Palestine Exploration Fund.
- De Luis M, Novak K, Raventos J, and Gricar J. 2011. Cambial activity, wood formation and sampling survival of Pinus halepensis exposed to different irrigation regimes. Forest Ecology and Management 262: 1630–1638.
- Esteban LG, Martin JA, de Palacios P, et al. 2010. Adaptive anatomy of Pinus halepensis trees from different Mediterranean environments in Spain. Trees 24: 19–30.
- Fady B. 2012. Biogeography of neutral genes and recent evolutionary history of pines in the Mediterranean Basin. Annals of Forest Science 69: 421–428.
- Fady B and Conord F. 2010. Macroecological patterns of species and genetic diversity in vascular plants of the Mediterranean basin. Diversity and Distribution 16: 53–64.
- Gómez A, Vendramin GG, González-martinez SC, and Alia R. 2005. Genetic diversity and differentiation of two Mediterranean pines (Pinus halepensis and Pinus pinaster Ait.) along a latitudinal cline using chloroplast microsatellite. Diversity and Distribution 11: 257–263.
- Grivet D, Sebastiani F, Gonzakez-Martinez SC, and Vendramine GG. 2009. Patterns of polymorphism resulting from long-range colonization in the Mediterranean conifer Aleppo pine. New Phytologist 184: 1016–1028.
- Grivet D, Sebastiani F, Alia R, et al. 2010. Molecular footprint of local adaptation in two Mediterranean conifers. Molecular and Biological. Evolution 28: 101–116.
- Grünwald C, Schiller G, and Conkle MT. 1986. Isoenzyme variation among native stands and plantations of Aleppo pine in Israel. Israel Journal of Botany 35: 161–174.
- Horowitz, A. 1979. The Quaternary of Israel. New York: Academic Press.
- INCO-DC. 2001. Global, physiological and molecular responses to climatic stresses of three Mediterranean conifers. International cooperation with developing countries (1994–1998). Contract No. ERBIC 18CT 970-2000. Final Report.
- Klein T, Di Matteo G, Rotenberg E, et al. 2012. Differential ecophysiological response of major Mediterranean pine species across a climatic gradient. Tree Physiology 33: 26–36.
- Knowles P and Grant MT. 1981. Genetic patterns associated with growth variability in Ponderosa pine. American Journal of Botany 68: 942–946.
- Knowles P and Mitton JB. 1980. Genetic heterozygosity and radial growth variability in Pinus contorta. Silvae Genetica 29: 114–118.
- Korol L and Schiller G. 1996. Relations between native Israeli and Jordanian Aleppo pine (Pinus halepensis) based on Allozyme anal: A note. Forest Genetics 3(4): 197–202.
- Korol L, Shklar G, and Schiller G. 2001. Diversity among Circum-Mediterranean populations of Aleppo pine and differentiation from Brutia pine in their isoenzymes: Additional results. Silvae Genetica 51(1): 35–41.
- Korol L, Shklar G, and Schiller G. 2002. Genetic variation within Pinus halepensis provenances growing in different microenvironments in Israel. Israel Journal of Plant Sciences 50: 135–143.
- Ledig FTh, Guries RP, and Bonefeld BA. 1983. The relation of growth to heterozygosity in Pitch pine. Evolution 37(6): 1227–1238.
- Maseyk SK. 2006. Ecophysiological and phenological aspects of Pinus halepensis in an arid-Mediterranean environment (Thesis). Rehovot: Weizmann Institute of Science.
- Mendel Z. 1984. Provenances as a factor in susceptibility of Pinus halepensis to Matsucoccus josephi (Homoptera: Margarodideae). Forest Ecology and Management 9: 259–266.
- Morandini R. 1976. Mediterranean conifers. Forest Genetic Resources, Information No. 5; Forestry Occasional Paper 1976/1. Rome: FAO.
- Nevo E. 1983. Population genetics and ecology: The interface. In: Bendall DS (Ed). Evolution from molecules to men. Cambridge (UK): Cambridge University Press.
- Nevo E. 1998. Molecular evolution and ecological stress at global, regional and local scales: The Israeli perspective. Journal of Experimental. Zoology 282: 95–119.
- Opal Y. 1978. Studies of morphological and chemical properties of Pinus halepensis in the Sha'ar Ha'gay Forest. Submitted to ORT, Technical College, Kefar-Sabba.
- [39] Oppenheimer HR. 1967. Mechanisms of drought resistance in conifers of the Mediterranean zone and in the arid West of the U.S.A. Part I: Physiological and anatomical investigations. Final report on project No. A10-FS7, Grant No. FG-Is-119. Rehovot: The Hebrew University of Jerusalem.
- Salim K, Naydenov KD, Benyounes H, et al. 2010. Genetic signals of ancient decline in Aleppo pine populations at the species' southwestern margins in the Mediterranean Basin. Hereditas 147: 165–175.
- Sathyan P, Newton RJ, and Loopstra CA. 2005. Genes induced by WSD are differentially expressed in two populations of Aleppo pine (Pinus halepensis). Tree Genetics & Genomes 1: 166–173.
- Schiller G and Grunwald C. 1986. Xylem resin monoterpene composition of Pinus halepensis in Israel. Israel Journal of Botany 35: 23–33.
- Schiller G and Grunwald C. 1987. Resin monoterpene in range-wide provenance trials of Pinus halepensis in Israel. Silvae Genetica 36: 109–114.
- Schiller G, Conkle MT, and Grunwald C. 1986. Local differentiation among Mediterranean populations of Aleppo pine in their isozymes. Silvae Genetica 35: 11–19.
- Schiller G and Waisel Y. 1989. Among-provenance variation in Pinus halepensis in Israel. Forest Ecology and Management 28: 141–151.
- Schiller G. 2000. Eco-physiology of Pinus halepensis and Pinus brutia Ten. In: Ne’eman G and Trabaud L (Eds). Ecology, biogeography and management of Pinus halepensis and P. brutia forest ecosystems in the Mediterranean basin. Leiden (The Netherlands): Backhuys Publishers.
- Schiller G, Korol L, and Shklar G. 2004. Habitat effects on adaptive genetic variation in Pinus halepensis provenances. Forest Genetics 11: 325–335.
- Schiller G and Atzmon N. 2009. Performance of Aleppo pine (Pinus halepensis) provenances grown at the edge of the Negev desert: A review. Journal of Arid Environments 73 (12): 1051–1057.
- Steinitz O. 2010. Gene flow between and within Aleppo pine (Pinus halepensis) populations (Thesis). Jerusalem: The Hebrew University of Jerusalem.
- Tognetti R, Michelozzi M, and Giovannelli A. 1997. Geographical variation in water relations, hydraulic architecture and terpen composition of Aleppo pine seedlings from Italian provenances. Tree Physiology 17: 241–250.
- Weinstein A. 1989. Provenance evaluation pf Pinus halepensis, brutia and P. eldarica in Israel. Forest Ecology and Management 26: 215–225.
- Weinstein-Evron M and Lev-Yadun S. 2000. Paleoecology of Pinus halepensis in Israel in the light of Palynological and Archeobotanical data. In: Ne’eman G and Trabaud L (Eds). Ecology, biogeography and management of Pinus halepensis and brutia forest ecosystems in the Mediterranean basin. Leiden (The Netherlands): Backhuys Publishers.
- Zohary M., Plant life of Palestine, Israel and Jordan. New York: The Ronald Press Company.











