
חזית המחקר
השפעת חוות הבודדים בנגב על המגוון הביולוגי בסביבתן
דניאל אורנשטיין הפקולטה לארכיטקטורה ובינוי ערים, הטכניון – מכון טכנולוגי לישראל ([email protected])
עמרת ליהוד התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב; המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגב
וואמויו מויטוי התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב; המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגב
עופר ארנון התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב
אלון טל המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגב ([email protected])

חוות בודדים שהוקמה על 'דרך היין' במרכז הנגב | צילום: דניאל אורנשטיין ©
אלי גרונר
מרכז מדע ים המלח והערבהדניאל אורנשטיין
הפקולטה לארכיטקטורה ובינוי ערים, הטכניון – מכון טכנולוגי לישראלעמרת ליהוד
התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב; המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגבוואמויו מויטוי
התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב; המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגבעופר ארנון
התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגבאלון טל
המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגבאלי גרונר
מרכז מדע ים המלח והערבהדניאל אורנשטיין
הפקולטה לארכיטקטורה ובינוי ערים, הטכניון – מכון טכנולוגי לישראלעמרת ליהוד
התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב; המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגבוואמויו מויטוי
התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגב; המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגבעופר ארנון
התכנית לתואר שני ללימודי מדבר, בי"ס כץ, אוניברסיטת בן-גוריון בנגבאלון טל
המחלקה לאקולוגיה מדברית, המכונים לחקר המדבר, אוניברסיטת בן-גוריון בנגבבמסגרת פרויקט "דרך היין" של המועצה האזורית רמת הנגב הוקמו בעשרים השנים האחרונות 27 חוות בודדים המתפרסות לאורך כביש 40 מאזור רביבים ועד למצפה רמון, יחד עם מספר חוות באזור ירוחם. תוספת של 27 יישובים ל-14 הקיימים באזור היא תוספת של 200% יישובים עם תוספת של כ-2% לאוכלוסייה, והיא עשויה לשנות את כל האקולוגיה ברמת הנגב. השפעתן של 15 חוות מ"דרך היין" על סביבתן נבדקה על פי מגוון ביולוגי של מכרסמים, זוחלים וחיפושיות. החוות נבדקו באביב ובקיץ במרחקים שונים מהחווה, החל מ-50 מטר וכלה ב-500 מטר מהחווה. כמו כן, נבדקו מספר אזורים ללא חוות (קבוצת ביקורת), והחוות עצמן נבחנו על פי העיסוקים בהן ושויכו על פי העיסוק העיקרי של החווה לקטגוריה הרלוונטית: תיירות, חקלאות בעלי חיים וחקלאות יבול.
בסך הכול נלכדו בשתי העונות 186 מכרסמים מארבעה מינים, 175 זוחלים מ-13 מינים ו-3,158 חיפושיות מ-26 מינים. הבדל מובהק בין מספר המכרסמים 500 מטר מהחווה למספרם בקרבת החווה ניכר רק בחוות העוסקות בתיירות. קבוצת החוות היחידה שהשפיעה על הימצאות זוחלים היא "חקלאות יבול". בקרבת חוות אלה היו יותר זוחלים מאשר במרחק של 500 מטר או בחוות אחרות. גם בכל הנוגע לחיפושיות רק לחוות חקלאות יבול יש השפעה על מספרן ועל הרכב החברה.
תוספת של חוות בודדים נוספות עשויה לשנות את הרכב החברה ואת המגוון הביולוגי באזור החוות. מספר המינים לאו דווקא ייגרע. הוא עשוי להישאר דומה או אפילו לעלות, אבל הרכב החברה עשוי להשתנות. במקרה כזה, מכיוון שהחוות מסודרות כמעט בקו ישר מצפון לדרום, עשוי להיווצר אזור חיץ שיהווה מעין חומה שתיצור שתי מערכות אקולוגיות בהר הנגב, מערבית ומזרחית.
על קצה המזלג
- ברחבי רמת הנגב הוקמו 27 חוות בודדים דלות אוכלוסייה, שעוסקים בהן בחקלאות (גידול יבול ו/או בעלי-חיים) ובתיירות.
- השפעתן הנוכחית של החוות על סביבתן נבדקה באמצעות מדידת המגוון הביולוגי של מכרסמים, חיפושיות וזוחלים במרחקים שונים מהחוות ובעונות שונות.
- תוצאות המחקר מראות כי אופי הפעילות בחווה באביב ובקיץ משפיע בצורה שונה על הקבוצות הטקסונומית השונות שנבדקו.
- פיזורן המרחבי של החוות עלול לחצות את המערכת האקולוגית של הר הנגב לשני אזורים מופרדים.
- הקמת חוות נוספות, כמתוכנן, עלולה לפגוע במגוון הביולוגי ובהרכב החברה סמוך להן.
המערכת
הלכה למעשה
מבוא
אף על פי שהנגב נחשף להשפעת אדם במשך אלפי שנים [6, 12], פעילות האדם במאה השנים האחרונות גבוהה יותר [29] והשפעתה על המגוון הביולוגי רבה יותר [35]. יישובים משפיעים מעבר לשטח שהם תופסים, דרך פעילות אדם סביב היישוב וניוד משאבים, אנשים, וכלי תחבורה ליישוב [18, 19]. השפעה זו עשויה להביא לאחידות של הנוף וליצור סכנה למינים מקומיים [9, 27].
באזור המדבר יכולה השפעת האדם להיות משמעותית ביותר בגלל תכונות המדבר. פעילות מטיילים ותיירים יכולה להשפיע על התנהגות, על מחזור החיים ועל תפוצה של בעלי חיים רבים, וכפועל יוצא מכך – על המגוון הביולוגי [14]. לחקלאות יכולה להיות השפעה קיצונית בגלל יצירת איים לחים בלב האזור הצחיח, בשל איי יצרנות גבוהה הנוצרים בעקבות הטיפולים הנלווים לחקלאות כגון קוטלי מזיקים, קוטלי עשבים ודשנים אחרים [34] ולאור סכנת המלחת הקרקע בגלל שימוש במים מטופלים [26]. גם הצטברות של פסולת יכולה לפגוע באקולוגיה של המדבר בגלל הפירוק האִטי של הפסולת, שקורה עקב לחות נמוכה [5, 28]. כמו כן, התיישבות אדם מביאה עִמה מפגעים נוספים מעבר לפסולת ולחקלאות, כגון חתולים וכלבים המסתובבים בשטח סביב ההתיישבות [7], חפירות, גדרות, ביוב ועוד. בעיה מרכזית נוספת הקשורה להתיישבות אדם באזורים עם הפרעה נמוכה היא יצירת קיטוע של בתי הגידול. הכנסת איים של הפרעות לשטח בלתי מופרע מביאה לירידה בגודל הממוצע של בית הגידול, להגדלת אזור הקצה ולבידוד בתי הגידול [16, 28]. מידת ההשפעה של יישובים על הנוף כולו תלויה במרחק שהיישובים משפיעים עליו: השפעה למרחק רב יכולה להביא ליצירת מסדרונות של הפרעה בלב המדבר. בסקר מומחים של המארג נבחנו האיומים המשפיעים על המגוון הביולוגי בהר הנגב, והאיום העיקרי זוהה כ"התיישבות הכוללת את החקלאות הסובבת אותה ותשתיות קוויות המלוות אותה" [1].
הבנתנו את ההשפעות האקולוגיות של פיתוח התיישבותי באזורים צחיחים לוקה בחסר (למעט השפעות על ציפורים). הגורמים העיקריים המביאים לירידה במגוון הביולוגי הם: א) איבוד בתי גידול; ב) התדרדרות בתי גידול כתוצאה משינויים מעשה ידי אדם; ג) כניסה של מינים לא מקומיים; ד) ציד וליקוט מיני בר [10]. מחקר על תגובת מגוון ביולוגי לפיתוח התיישבותי באזורים צחיחים מעיד על שיפור אצל אוכלוסיות מינים כוללנים (generalist) וחובבי לחות ועל הרעה אצל מינים אנדמיים או כאלה המתמחים במדבר [21]. תפוצת ציפורי שיר, למשל, הגיבה לפיתוח התיישבותי בהתאם לשתי קבוצות ברורות: א) מינים חובבי אדם, שצפיפותם עלתה בנוכחות אדם; ב) מינים רגישים לנוכחות אדם, שצפיפותם ירדה עם ההתיישבות. כדי להשאיר מרחבים גדולים ככל האפשר ללא נוכחות אדם הומלץ על בנייה מרוכזת [22]. כמו כן, Trammel ואחרים [33] מצאו מספר גבוה של מיני ציפורים באזור פיתוח עירוני בזכות מסדרון ירוק.

במועצה האזורית רמת הנגב התפתחה בשנים האחרונות צורת התיישבות חדשה המבוססת על משפחה בודדת הגרה באזור מבודד יחסית מיישובים אחרים, ונקראת "חוות בודדים" [2, 24]. חוות בודדים קיימות גם באזורים אחרים בארץ. ב-1990 התקבלה החלטה מספר 443 של מִנהל מקרקעי ישראל שנועדה להסדיר מסירת קרקע למשפחות יחידים למטרות התיישבות מחוץ ליישובים [2]. יש ברמת הנגב 33 חוות בודדים, מתוכן 27 חוות על "דרך היין". רובן הוקמו ב-15 השנים האחרונות על שטח ממוצע של 2,000 דונם לחווה. החוות נמצאות בקרבת כביש 40 ויוצרות מסדרון בצורת Y מאזור קיבוץ רביבים וירוחם ועד למצפה רמון (איור 1). החוות יוצרות קו של התיישבות חקלאית/תיירותית במרכז הנגב, ומחלקות את הנגב המרכזי לאורך. אם לחוות אלה השפעה על האזור סביב החווה, הרי שעשויה להיות סכנה של יצירת חומה של השפעת האדם המחלקת את הנגב המרכזי לנגב מערבי ולנגב מזרחי ויוצרת קיטוע. אם מגמת ההתיישבות בנגב תמשיך באותה מגמה (כ-15 חוות בעשור), הרי שיש חשיבות רבה לבחינת מידת ההשפעה של החוות על הקיטוע ועל המגוון הביולוגי סביבן. רצוי לבחון אם לאופי החווה – עיסוק חקלאי או תיירותי – יש השפעה על הסביבה של החווה.

איור 1
חוות הבודדים שנדגמו על פי העיסוק העיקרי בחווה
אנחנו תופסים שתי תכונות של פיתוח כחשובות ביחס להשפעות אקולוגיות: צפיפות אוכלוסין ושימושי קרקע של התיישבות. ביחס לצפיפות האוכלוסין, שיעור ההשפעה המרחבית של שינוי קרקע של כל קהילה (כגון קיבוץ, עיר, חווה) יכול להיות זהה מבחינת השטח, אולם יש לבחון זאת כהשפעה ביחס לכל ראש. דבר זה חשוב במיוחד, מכיוון שהנגב נחשב כשסתום שחרור ללחץ הדמוגרפי של מרכז הארץ [8]. התכונה השנייה, שימושי קרקע הקשורים לפיתוח התיישבותי, כגון חקלאות ומרעה, חשובה בגלל שהיא קובעת את שיעור ההשפעה המרחבית של האדם ואת עוצמתה מעבר למדרך האקולוגי ביישובים עצמם. באזורים מדבריים תורמת החקלאות להמלחה ולסחף של קרקע, מצריכה שימוש בכמות מים לא פרופורציונית לרווח הכלכלי, ונחשבת גורם מכריע במִדבור [26]. השפעת רעייה (מבויתת) על מגוון ביולוגי תלויה בעוצמת הרעייה. בצפון הנגב קיימת רעייה אלפי שנים, וחברת הצמחים התפתחה תחת לחץ זה ובמקרים מסוימים היא אף תלויה בלחץ מתון לשמירה על הרכבה [25]. Olsvig-Whittaker ואחרים [23] הסיקו שפיתוח עירוני וחקלאות, ולא רעייה, הם האיומים העיקריים על השטחים המדבריים.
בחנו את השפעת החוות על המגוון הביולוגי של שלוש קבוצות טקסונומיות נבחרות סביב החוות ולפי התעסוקה העיקרית בחוות, במטרה לבחון כיצד עשוי המשך ההתיישבות בנגב להשפיע על הסביבה בו.
שיטות
חוות
15 חוות נבחרו על פי סקר מוקדם שנערך במטרה לבדוק אילו חוות מתאימות לבחינת שאלות המחקר. במסגרת המשאבים המצומצמים שלנו העדפנו לבדוק מספר רב ככל האפשר של חוות על פני דגימה במספר רב של ימים אך במספר חוות קטן יותר, מכיוון שהחזרות האמִתיות לשאלת המחקר הן החוות עצמן. התנאים שנבחנו הם מרחק ממבנים ומגורמים אחרים בקרבת החוות שעשויים להשפיע על הסביבה. העדפנו לבחון חוות שאינן בקרבת מִכרות, בסיסי צבא והפרעות אחרות, כדי לבודד את השפעתן, שניתנת למדידה בקלות יחסית. כמו כן הובאו בחשבון הגישה לחווה, שיתוף הפעולה עם החוואים והאפשרות למצוא שטח ביקורת בקרבתה.
סוג פעילות
שייכנו כל חווה לתחום העיסוק העיקרי שלה. תחומי העיסוק הם: חוות גידולי מזרע (להלן חקלאות יבול), הכוללת גידול צמחים; חקלאות חי (להלן חקלאות בעלי חיים), הכוללת גידול בעלי חיים כגון עִזים, כבשים ופרות; חוות תיירות, הכוללות בתי אירוח, מסעדות ופעילויות אחרת.
חלקות
בכל חווה נבחרו 12 חלקות, בגודל של 400 מ"ר כל אחת, שפוזרו באופן שווה על שלושה מעגלים ברדיוסים של 50, 100 ו-500 מטר. המרחקים נבחרו כך שיהיה ניתן לבחון את השפעת החווה באזור שקרוב אליה ובמרחק רב ככל האפשר. מרחק של 500 מטר נבחר כאזור שמרוחק דיו מהחווה כדי לא להיות מושפע מגורמים אנתרופוגניים, ועם זאת, הרי הוא בית גידול דומה לאזור החווה. כמו כן, הוחלט להכניס חלקות ביקורת כלליות שאינן קשורות לחווה מסוימת, שנמצאות בבית גידול דומה ומרוחקות מכל השפעות אדם. לשם כך, נבחרו חלקות במרחק של 2 ק"מ מכל התיישבות בבית גידול סלעי. בקיץ נדגמו 12 חלקות ביקורת באזור עבדת, ובאביב נדגמו 48 חלקות ביקורת בארבעה אתרים שונים: צפונית לצומת חלוקים, דרומית לצומת חלוקים, דרום-מערבית לכלא נפחא ומזרחית לכלא נפחא. החוות נבדקו בקיץ 2009 (אוגוסט–אוקטובר) ובאביב 2010 (מרץ–מאי), והניתוח נעשה לכל עונה בנפרד.
לכידות
הלכידות במחקר זה נעשו באופן שאיננו הורג את בעלי החיים או פוגע בהם. כל המלכודות היו יבשות וללא רעל, וכמעט כל בעלי החיים זוהו מיד ושוחררו בקרבת המלכודת.
לכידת מכרסמים – השתמשנו במלכודות שרמן שהונחו בבתי גידול סלעיים סביב החווה. לאחר השוואה בין בתי גידול של ואדיות עם כיסוי צומח רב לבין אזורים סלעיים, רוכז מאמץ הדגימה באזור הסלעי, שם נצפו יותר לכידות. המלכודות הופעלו 3-2 לילות בכל אחת מהעונות שנבדקו. המלכודות הונחו אחה"צ ונבדקו השכם בבוקר שלמחרת. באזור כל חווה נדגמו 12 חלקות כשבכל חלקה 10 מלכודות במרחק של 10 מטר אחת מהשנייה. לכידה חוזרת נבדקה על-ידי סימון בטוש בצד הגחוני (ventral), רק כדי לאמת את הקשר בין כמות הלכידות לצפיפות, אך המשתנה שמוצג הוא מספר הלכידות ולא גודל האוכלוסייה. הלכידות לא נעשו בלילות ירח מלא [11, 14], בלילות שמזג האוויר בהם היה גשום או בסופות חול שעלולות לסגור את המלכודות ולנטרלן.
לכידות זוחלים – כדי לתפוס בעלי חיים קטנים, שוכני קרקע, הונחו מלכודות נפילה יבשות בחלקות סביב החוות. המלכודות היו דליים בעלי קוטר של 36 ס"מ ועומק של 18 ס"מ, שפתחם בגובה פני הקרקע. הם כוסו באבן גדולה שכיסתה רק חלק מהפתח ואפשרה לבעלי חיים ליפול לדלי. בכל חלקה היו חמש מלכודות נפילה, כך שבכל חווה בכל אחד משלושת המרחקים היו 20 מלכודות בארבע חלקות. המלכודות נבדקו פעם בשבוע במשך חודשיים בקיץ (יולי–אוגוסט) ובאביב (אפריל–מאי). בעלי החיים זוהו במקום ושוחררו. לכידת חיפושיות – לכידת החיפושיות נעשתה באמצעות אותן מלכודות (דליים) ששימשו ללכידת הזוחלים, באותם מקומות ובאותם מועדים. זיהוי החיפושיות נעשה בשטח, ובמקרה של ספק הן הובאו למעבדה, זוהו (על-ידי המחבר הראשון) ושוחררו.
צפיפות/פעילות בעלי החיים
המשתנה שהושווה בין החוות והחלקות הוא פעילות בעלי חיים. משתנה זה של צפיפות הלכידוֹת כולל את צפיפות בעלי החיים באזור ואת מידת פעילותם בשטח. משתנה זה נקרא בספרות 'פעילוּת' [13], ונלקח כמדד עקיף לגודל האוכלוסייה בשטח ולמידת הפעילות שלה. הוא יעיל כאשר נעשה מחקר המשווה בין מקומות שונים [9] ואינו מצביע על הצפיפות המוחלטת של האוכלוסייה. כדי לעשות כיול חולק מספר הפרטים במאמץ הדגימה על פי המשוואה הבאה:
משוואה 1: A = I / t*d
כאשר A=פעילות, I=מספר פרטים שנלכדו, t=מספר המלכודות ו-d=מספר הימים שהמלכודות היו פתוחות.
ניתוח נתונים
בחנו את השפעת סוג החווה והמרחק מהחווה על מספר בעלי החיים שנלכדו עבור כל הקבוצות הטקסונומיות. עבור קבוצות עם מספר מינים רב (חיפושיות וזוחלים) בחנו גם את ההשפעה על מספר המינים והרכבם. מספר הלכידות והמינים של כל הקבוצות הטקסונומיות עברו טרנספורמציה לוגריתמית ונבדקו לנורמליות באמצעות מבחן קולמגורוב-סמירנוב.
מכיוון שהשערת המחקר הייתה שהחוות משנות את הרכב החברה, ולאו דווקא מעלות או מורידות את מגוון המינים, בדקנו את מידת הדמיון בין החוות עבור החיפושיות. הדמיון בין האתרים נעשה בעזרת ניתוח רב-משתנים הנקרא אורדינציה [31]. נעשה ניתוח (PCA (Principal Component Analysis המארגן את האתרים על פי מידת הדמיון שלהם בהרכב החברה, ובכלל זה צפיפות אוכלוסיות של מינים דומיננטיים. ה-PCA יוצר איור המראה לא רק את הקִרבה בין אתרים דומים, אלא גם את הקישור של האתרים למינים הדומיננטיים בהם [31]. כמו כן נעשה ניתוח אורדינציות ישיר RDA Redundancy Analysis המסתמך על אורדינציה לינארית ועל סימולציית מונטה קרלו עם 9999 הרצות. באורדינציה הישירה בדקנו את ההשערה שסוג החווה משפיע על הרכב החברה. כל ניתוחי האורדינציות נעשו בתכנת [31] CANOCO.
תוצאות
מכרסמים
בסך הכול היו 278 לכידות של 186 פרטים מארבעה מינים. רוב הפרטים (132) היו מסוג גרביל סלעים (Gerbillus dasyurus), ו-45 מסוג קוצן מצוי (Acomys cahirinus). הלכידות החוזרות של המכרסמים נעשו כמעט תמיד באתר הלכידה המקורי. רק בשני מקרים אירעו לכידות חוזרות במרחק של כ-50 מטר מאתר הלכידה המקורי.
צפיפות המכרסמים שנתפסו באביב ליד החוות הייתה נמוכה מצפיפותם בחלקות הביקורת (F=3.432, p=0.0187). לאופי העיסוק בחווה הייתה השפעה על מספר המכרסמים: בקרבת חוות תיירות (t=2.803, DF=148, p=0.0057) וחקלאות בעלי חיים (t=2.34, DF=148, p=0.0206) נתפסו פחות מכרסמים מאשר בחלקות הביקורת (איור 2). לעומת זאת, בקרבת חוות חקלאות יבול נתפסו מכרסמים במספר שאינו שונה באופן מובהק מזה שבחלקות הביקורת. באופן כללי לא נצפתה השפעה של המרחק מהחוות על מספר המכרסמים שנתפס, למעט חוות חקלאות בעלי חיים: מספר המכרסמים בחלקות הנמצאות 500 מטר מחוות חקלאות בעלי חיים, גבוה ממספר המכרסמים בחלקות המרוחקות 100 מטר מהחוות (t=2.405, DF=27, p=0.023). בקיץ, בחוות תיירות, תועדו יותר גרבילי סלעים במרחק של כ-500 מטר מאשר ב-100 וב-50 מטר (F(2,21)=6.48, p=0.0064). עיקר ההבדלים בין המכרסמים נובע מהקוצנים: מספר הגרבילים בחוות השונות היה דומה (קיץ F=0.4828, p=0.69; אביב F=1.696, p=0.17), בעוד שמספר הקוצנים בחוות היה שונה (קיץ F=5.766, p=0.001; אביב F=5.523, p=0.0013), ושיעור לכידתם בחלקות הביקורת היה גבוה מזה שבחוות.
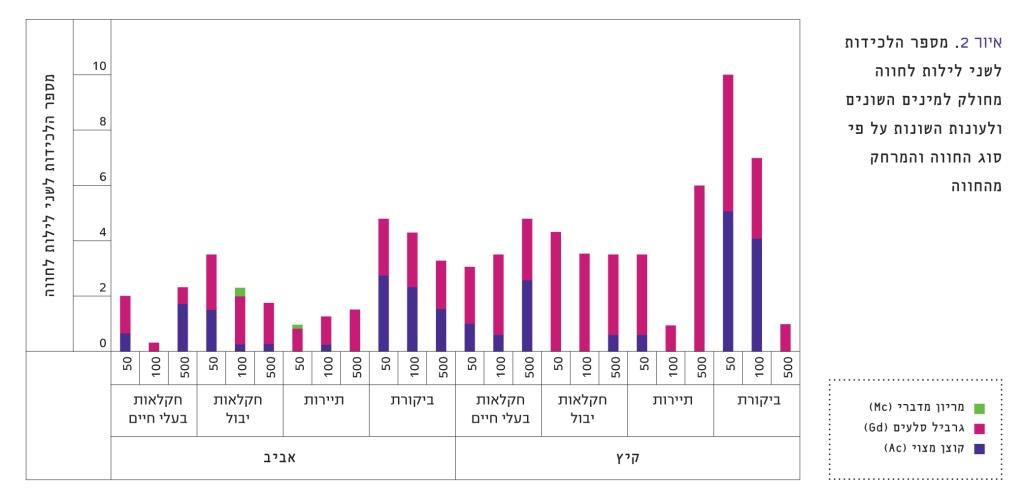
איור 2
מספר הלכידות לשני לילות לחווה מחולק למינים השונים ולעונות השונות על פי סוג החווה והמרחק מהחווה
בקיץ לא השפיע אופי החווה על מספר המכרסמים. ההשפעה היחידה שנצפתה הייתה במרחק מחוות תיירות: בקיץ היה מספר המכרסמים שנתפסו במרחק 500 מטר מהחוות גדול יותר ממספר המכרסמים בחלקות של 50 ו-100 מטר (מבחן שונות חד-כיוונית F=6.472, p=0.0065). באופן כללי, היו יותר לכידות בחוות בקיץ ביחס למאמץ הדגימה מאשר באביב (איור 2). איור זה מתאר את מספר הלכידות לחווה לפי מינים, עונות, סוג החווה והמרחק מהחווה (עבור שני לילות).
זוחלים
בסך הכול נלכדו 175 זוחלים מ-13 מינים. בקיץ היה מספר הזוחלים גבוה יותר בקרבת חוות חקלאות יבול מאשר בסוגי החוות האחרות. במבחן אנליזה של שונות דו-גורמית (מרחק מהחווה וסוג החווה) על מספר הזוחלים נמצא כי המרחק איננו מובהק (F=0.82, p=0.45), סוג החווה מובהק (F=31.61, p=0.043) ואילו האינטראקציה בין החווה למרחק אינה מובהקת (F=1.95, p=0.12).
מספר הזוחלים היה גבוה בקרבת חוות חקלאות יבול גם בחלקות המרוחקות יותר (F=7.99, p=0.006), ובעיקר נכון הדבר לגבי ישימונית מצויה (Stenodactylus sthenodactylus) ומדברית עינונית (Mesalina guttulata). מינים אלה נמצאו בצפיפות גבוהה יותר בקרבת החוות, וצפיפותם יורדת עם המרחק מהחווה. מגמה זו מתהפכת באביב, שבו מספר הזוחלים יורד במעט עם ההתקרבות לחוות. במבחן אנליזה של שונות דו-גורמית (עונה וסוג החווה) על מספר הזוחלים נמצא כי העונה מובהקת (F=8.71, p=0.005), המרחק איננו מובהק (F=2.21, p=0.12), ואילו האינטראקציה בין המרחק לעונה מובהקת (F=3.37, p=0.042). אין השפעה למרחק או לסוג החווה על צפיפות הזוחלים או על מספר המינים באביב.
חיפושיות
בסך הכול נלכדו 3,158 חיפושיות מ-26 מינים. בחינת השפעת המרחק מהחוות וסוג החוות על מספר החיפושיות בקיץ העלתה כי רק סוג החוות השפיע. הנתונים הראו התפלגות נורמלית לאחר טרנספורמציה לוגריתמית. במבחן אנליזה של שונות דו-גורמית (מרחק מהחווה וסוג החווה) על מספר החיפושיות נמצא כי המרחק איננו מובהק (F=1.27, p=0.29), סוג החווה מובהק (F=31.61, P<0.001) ואילו האינטראקציה בין החווה למרחק גבולית (F=2.43, P=0.07). מספר הפרטים בקרבת חוות חקלאות יבול היה נמוך באופן מובהק ממספר הפרטים בקרבת חוות חקלאות בעלי חיים וחוות תיירות. האינטראקציה בין סוג החווה למרחק הייתה גבולית (F=2.42, P=0.07). גם מספר המינים דומה במרחקים שונים בקיץ בקרבת חוות תיירות וחוות חקלאות בעלי חיים, אך נמוך יותר בקרבת חוות חקלאות יבול. באביב, לעומת זאת, מספר החיפושיות שנלכדו בקרבת חוות חקלאות יבול איננו שונה מחוות אחרות ואף גבוה במעט (איור 3). ישנה אינטראקציה מובהקת בין סוג החווה לעונה לגבי מספר החיפושיות שנלכדו. במבחן אנליזה של שונות דו-גורמית (עונה וסוג החווה) על מספר החיפושיות נמצא כי העונה מובהקת (F=52.73, P<0.001), סוג החווה מובהק (F=6.15, P=0.003) וגם האינטראקציה בין החווה לעונה מובהקת (F=17.08, P<0.001).
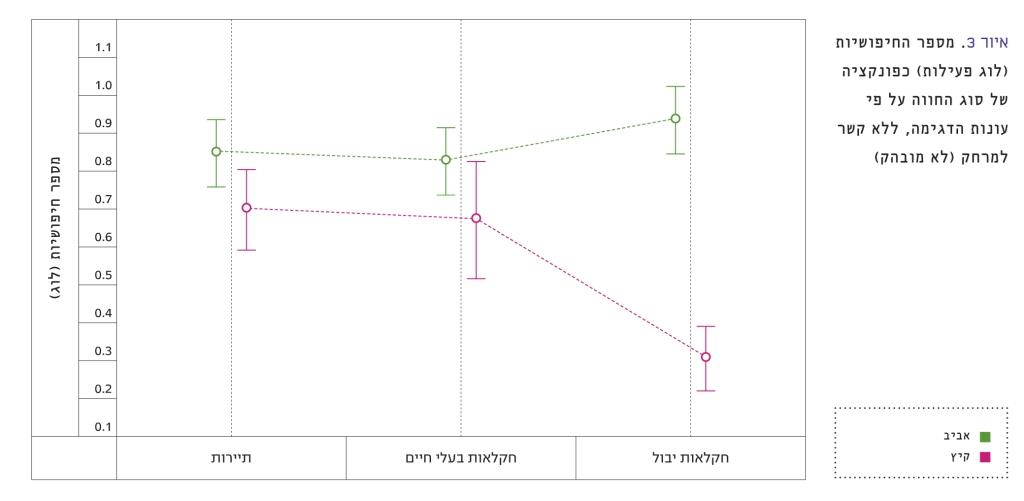
איור 3
מספר החיפושיות (לוג פעילות) כפונקציה של סוג החווה על פי עונות הדגימה, ללא קשר למרחק (לא מובהק)
הרכב מאספי החיפושיות שונה בין סוגי החוות השונות באביב (איור 4, RDA: P=0.0240, F=3.53) ובקיץ (RDA: P=0.0020, F= 10.06). איור 4 מראה את הקשר בין סוג החוות למינים מסוימים: מין שנמצא סמוך לסוג חווה מעיד על קשר ביניהם. סוגי תעסוקה שונים הנמצאים במרחק גדול מעידים על השפעה של סוג החווה על הרכב החברה. באביב חוות חקלאות בעלי החיים שונות מהחוות האחרות (איור 4א), בעוד שבקיץ חוות חקלאות היבול שונות מהחוות האחרות (איור 4ב).
ישנם מספר מינים נפוצים ביותר באביב (איור 4א) כמו אירודית (Erodius reichei) ומזוסטנה (Mesostena angustata), הנמצאים בקרבת חוות תיירות, ובצפיפות נמוכה יותר בקרבת חוות חקלאות בעלי חיים. בקיץ ישנם מינים, כמו פצירנית (Trachiderma philistini), פימליה גדולה (Pimelia grandis), אקיס (Akis goryi) ובלפס הרכסים (Blaps sulcata), הנמצאים בצפיפויות גדולות בקרבת חוות חקלאות בעלי חיים וחוות תיירות ובצפיפויות נמוכות בקרבת חוות חקלאות יבול (איור 4ב).

איור 4
הרכב מאסף החיפושיות באביב (א) ובקיץ (ב) על פי אורדינציות PCA
דיון
תוצאות המחקר מעידות שקבוצות שונות מגיבות באופן שונה לחוות ובאופן שונה מאד בין העונות. הפעילות בחווה קובעת את ההשפעה של החווה על הסביבה. באופן כללי, באזור החוות העוסקות בתיירות יש פחות מכרסמים מאשר בחוות האחרות. יש ירידה במספר החיפושיות בקיץ ככל שמתקרבים לחוות חקלאות. כשמתקרבים לחוות חקלאות יבול יורד מספר החיפושיות, בעיקר ממינים נפוצים ממשפחת השחרוריות כגון בלפס, פימליה ופצירנית. מספר הזוחלים עולה ככל שמתקרבים לחוות, ובעיקר נכון הדבר לגבי חוות חקלאות יבול בקיץ. באביב הנתונים הטרוגניים יותר, עם נטייה לירידה קלה במספר הזוחלים ככל שמתרחקים מהחוות.
ההיסטוריה של אזור הר הנגב רצופה בסוגים שונים של התיישבויות מאז התקופה הפלאוליטית, לפני למעלה מעשרת אלפים שנה [6] וכוללת שגשוג של ממלכות שונות. התיישבויות אלה, שכללו חקלאות ומרעה, התבססו על ניצול נכון של המשאב המגביל העיקרי במדבר, המים, בדרכים שונות, כדוגמת טרסות ושיחים. צורת חיים זו אפשרה קיום מאוזן עם הסביבה [12]. מאז קום המדינה החלה התיישבות מדגם שונה, המתבססת על מקורות מים מחוץ לאזור. ההתיישבות עד לעשור האחרונה כללה שתי עיירות וכעשרה יישובים קטנים. בעשור האחרון הושלש מספר היישובים על-ידי הכנסת חוות הבודדים. נוצר מצב חדש בהר הנגב – יש מספר רב מאוד של יישובים המקבלים מים מחוץ לאזור, אם כי גודל האוכלוסייה עדיין קטן באופן יחסי. התוצאה היא רצף של איים ירוקים בלב המדבר. חוות הבודדים עשויות לפגוע ברצף של מערכות אקולוגיות ומגוון ביולוגי בהר הנגב [4]. בחלק מהנתונים שלנו נראה כי עצם קיום החווה משנה את הרכב החברה בהשוואה לקבוצת הביקורת, כי אופי החווה משפיע, וכי גם במרחק של 500 מטר עדיין ניכרים השינויים הקשורים לחווה עצמה. אם החווה משפיעה על הרכב החברה למרחק היפותטי ברדיוס 1 ק"מ, עשוי להיווצר בהר הנגב מסדרון של מערכת אקולוגית עם חברה שונה (איור 5). מסדרון זה יכול לאפשר למינים שונים לחדור אל תוך המדבר הקיצון וכן להיות חיץ וליצור שתי מערכות אקולוגיות בהר הנגב: מערבית ומזרחית, שהמסדרון שהוזכר לעיל חוצץ בין מינים ואוכלוסיות משתי המערכות האקולוגיות. השטח הבלתי מופר יהפוך לקטוע ביותר.
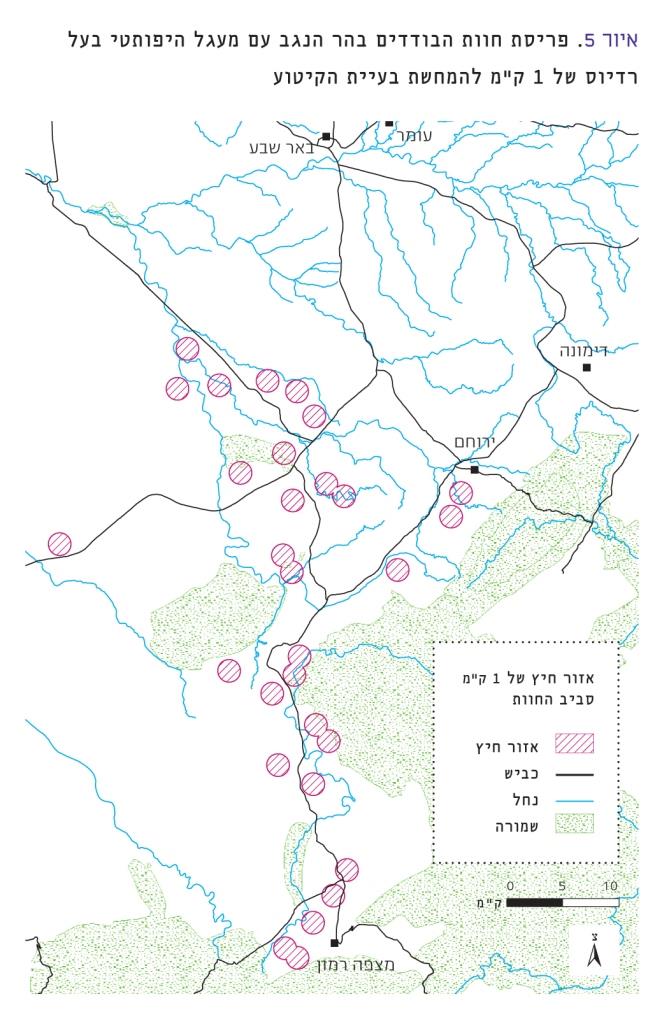
איור 5
פריסת חוות הבודדים בהר הנגב עם מעגל היפותטי בעל רדיוס של 1 ק"מ להמחשת בעיית הקיטוע
מן המחקר מתברר שלחוות אין השפעה חד-כיוונית בשטח שסביבן. לא נצפתה בשום קבוצה ירידה מונוטונית של מספר הפרטים או פגיעה במגוון הביולוגי כתלות במרחק בכל העונות. באופן כללי, במרחקים שבדקנו לא נצפה הבדל משמעותי שנובע ממידת הקִרבה לחווה. רוב ההבדלים שמצאנו היו בין חלקות הביקורת לחוות ובין סוגי החוות. החוות נמצאות בשטח הטרוגני, כל חווה נמצאת בתוואי שטח מעט שונה, ולכן ההשוואה ביניהן לא פשוטה. שום קטגוריה לא בולטת בהשפעות השליליות שלה. בעוד שחוות תיירות וחוות חקלאות בעלי חיים כנראה מורידות את צפיפות המכרסמים, חוות חקלאות יבול מעלות את מספר הזוחלים ומורידות את צפיפותן של חלק מהחיפושיות. ברור שישנם מינים רבים אחרים המושפעים מהחוות שלא בדקנו, כולל מינים נדירים, מינים מתפרצים הנלווים לאדם ואחרים. באותה מידה שקבוצות שונות הגיבו באופן דיפרנציאלי לחוות, כך גם צפוי שההשפעה של החוות על כלל המגוון הביולוגי תהיה דיפרנציאלית.
הנגב מכיל עירוב של פיתוח התיישבותי ובו מושבים חקלאיים, קיבוצים, חוות בודדים, עיירות פיתוח ופזורה בדואית. ניתן לאפיין כל אחד מאלה בעזרת הגורמים החשובים המשפיעים על אקולוגיה: צפיפות אוכלוסין ושימושי קרקע. חוות הבודדים הן סוג ההתיישבות שצפיפות האוכלוסין בו היא הנמוכה ביותר – משפחה אחת בשטח החווה. להבדיל, בקיבוץ שדה בוקר מתגוררים כ-370 תושבים ובמדרשת בן-גוריון כ-1,200 תושבים [3]. לכן, כל השפעה הנצפית מחוות בודדים היא השפעה של משפחה בודדת אחת. אין מחקר זה טוען שהחוות משפיעות באופן שונה על הסביבה מאשר קיבוצים, מושבים, הפזורה הבדואית ואף עיירות באזור. ההבדל היחיד הוא מספר החוות ביחס לגודל האוכלוסייה הנתמכת. 27 משפחות שיתגוררו ביישוב אחד ישפיעו פחות על הסביבה מאשר 27 חוות.
אנו מתייחסים לחוות הבודדים כאל חלק מהפרדיגמה של הפיתוח ההתיישבותי, שגורמת לזחילה עירונית. סוג זה של התיישבות משמעותי להשפעות אקולוגיות משלוש סיבות: ראשית, גם אם לחוות השפעות דומות לאלה של סוגי ההתיישבות האחרות ששטחן דומה, ההשפעה לכל אדם גבוהה ביותר. שנית, להבדיל מיישוב עירוני כמו ירוחם, מצפה רמון ובאר שבע, שימוש הקרקע העיקרי הוא חקלאות (כמו הקיבוצים והמושבים) שהוא המשפיע ביותר [26]. ושלישית, החוות ממוקמות באזורים רגישים אקולוגית, ובמקרים רבים אף סמוך לשמורות טבע [15]. מיקום זה עשוי להביא לכך שפעילות חוות בודדים תשפיע על מינים בשמורות טבע, שקודם לכן היו רחוקים ממקומות יישוב.
לא מפתיע שהתוצאות שלנו רק מרמזות על השפעה ואינן החלטיות. תגובות אקולוגיות לשינויים בשימושי קרקע מתבטאות בתהליכים ארוכי טווח שמצריכים ניטור ארוך טווח שעשוי להימשך גם עשורים [15]. תוצאות המחקר שלנו מרמזות על שינויים המתרחשים כתוצאה מהקמת החוות, ולכן יש להמשיך לעקוב אחר שינויים אלה לאורך זמן. לפי פרדיגמת שמירת הטבע [20, 32], לפיתוח התיישבותי כגון זה של חוות הבודדים, יש השפעה רבה בעיצוב המגוון הביולוגי של מרכז הנגב. שינויים בהרכב החברה יכולים להוביל להיכחדות של אוכלוסיות ולירידה בתפקוד של המערכות האקולוגיות בנתינת שירותים אקולוגים לאדם [17]. תוצאות מחקר זה מחזקות את הצורך בבחינת דרכי התיישבות צמודות דופן אל מול הקמת יישובים נוספים, כמו גם בשקילת מספר היישובים כמשתנה חשוב בבחינת השפעת האדם על הסביבה בנגב.
- ברחבי רמת הנגב הוקמו 27 חוות בודדים דלות אוכלוסייה, שעוסקים בהן בחקלאות (גידול יבול ו/או בעלי-חיים) ובתיירות.
- השפעתן הנוכחית של החוות על סביבתן נבדקה באמצעות מדידת המגוון הביולוגי של מכרסמים, חיפושיות וזוחלים במרחקים שונים מהחוות ובעונות שונות.
- תוצאות המחקר מראות כי אופי הפעילות בחווה באביב ובקיץ משפיע בצורה שונה על הקבוצות הטקסונומית השונות שנבדקו.
- פיזורן המרחבי של החוות עלול לחצות את המערכת האקולוגית של הר הנגב לשני אזורים מופרדים.
- הקמת חוות נוספות, כמתוכנן, עלולה לפגוע במגוון הביולוגי ובהרכב החברה סמוך להן.
המערכת
מקורות
- ברג נ ופרבולוצקי א. 2011. התכנית הלאומית לניטור המגוון הביולוגי בשטחים פתוחים בישראל. ירושלים: המארג.
- האן א, שגיא י, בורל ר ודרום י. 2003. התיישבות חדשה: יישובים חדשים והתיישבות בודדים – עמדת החברה להגנת הטבע. תל-אביב: החברה להגנת הטבע.
- הלשכה המרכזית לסטטיסטיקה. 2008. מפקד אוכלוסין. ירושלים: משרד הפנים.
- הבלנה ד. 2005. השפעות אקולוגיות של התיישבות חדשה בנגב. ירושלים: רשות הטבע והגנים.
- Abeliovich A and Ronen Z. 1997. Pollution and bioremediation of soils in arid zones. In: Irvine RL and Sikdar SK (Eds). Biodegradation technology developments: Principles and practice. Basel, Switzerland: CRC Press.
- Avni Y. 2005. Gully incision as a key factor in desertification in an arid environment, the Negev highlands, Israel. Catena 63(2–3): 185–220.
- Brinkner-Braun I, Geffen E, and Yom-Tov Y. 2007. The domestic cat as predator of Israeli wildlife. Israel Journal of Ecology and Evolution 53(2): 129–142.
- Cohen SB. 2000. The polyurban frontier in post-industrial Israel. In: Lithwick H and Gradus Y (Eds). Developing frontier cities: Global perspectives – regional contexts. Dordrecht: Kluwer Academic Publishers.
- Crooks KR, Suarez AV, and Bolger DT. 2004. Avian assemblages along a gradient of urbanization in a highly fragmented landscape. Biological Conservation 115(3): 451–462.
- Darkoh MBK. 2002. Regional perspectives on agriculture and biodiversity in the drylands of Africa. Journal of Arid Environments 54: 261–279.
- Eilam D. 2004. Locomotor activity in common spiny mice (Acomys cahirinus): The effect of light and environmental complexity. BMC Ecology 4(1): 16.
- Evenari M, Shanan L, and Tadmor N. 1982. The Negev – The challenge of the desert, 2nd ed. Cambridge (MA): Harvard University Press.
- Groner E and Ayal Y. 2001. The interaction between bird predation and plant cover in determining habitat occupancy of darkling beetles. Oikos 93(1): 22–31.
- Gutzwiller KJ. 1995. Recreational disturbance and wildlife communities. In: Knight RL and Gutzwiller KJ (Eds). Wildlife and recreationists: Coexistence through management and research. Washington (DC): Island Press.
- Hansen AJ, Knight RL, Marzluff JM, et al. 2005. Effects of exurban development on biodiversity: Patterns, mechanisms, and research needs. Ecological Applications 15: 1893–1905.
- Kotler BP, Ayal Y, and Subach A. 1994. Effects of predatory risk and resource renewal on the timing of foraging activity in a gerbil community. Oecologia 100(4): 391–396.
- Luck GW, Daily GC, and Ehrlich PR. 2003. Population diversity and ecosystem services. Trends in Ecology and Evolution 18(7): 331–336.
- Luck GW, Ricketts TH, Daily GC, and Imhoff M. 2004. Alleviating spatial conflict between people and biodiversity. Proceedings of the National Academy of Sciences of the United States of America 101(1): 182–186.
- Marzluff JM and Neatherlin E. 2006. Corvid response to human settlements and campgrounds: Causes, consequences, and challenges for conservation. Biological Conservation 130(2): 301–314.
- Meffe GK and Carroll CR. 1994. Principles of conservation biology. Sunderland (MA): Sinauer Associates, Inc.
- Miller JR, Wiens JA, Thompson Hobbs N, and Theobald DM. 2003. Effects of human settlement on bird communities in lowland riparian areas of Colorado. Ecological Applications 13: 1041–1059.
- Odell EA and Knight RL. 2001. Songbird and medium-sized mammal communities associated with exurban development in Pitkin County, Colorado. Conservation Biology 15(4): 1143–1150.
- Olsvig-Whittaker L, Frankenberg E, Perevolotsky A, and Ungar ED. 2006. Grazing, overgrazing and conservation: Changing concepts and practices in the Negev rangelands. Sécheresse 17(1/2): 195–199.
- Orenstein DE, Jiang L, and Hamburg SP. 2011. An elephant in the planning room: Political-demography and its influence on sustainable land use planning in drylands. Journal of Arid Environments 75(6): 596–611.
- Perevolotsky A and Seligman NG. 1998. Role of grazing in Mediterranean rangeland ecosystems. BioScience 48(2): 1007–1017.
- Portnov B and Safriel U. 2004. Combating desertification in the Negev: Dryland agriculture vs. dryland urbanization. Journal of Arid Environments 56(4): 659–680.
- Ro TH and Hong SK. 2008. Landscape ecology for biodiversity – scaling up. In: Hong SK, Nakagoshi N, Fu BJ, and Morimoto Y (Eds). Landscape ecological applications in man-influenced areas – linking man and nature systems. The Netherlands: Springer.
- Sanderson EW, Jaiteh M, Levy MA, et al. 2002. The human footprint and the last of the wild. BioScience 52(10): 891–904.
- Shenbrot G and Krasnov B. 2004. Temporal dynamics in spatial organization of a rodent community in the Negev Highlands (Israel). Journal of Zoology 263: 207–218.
- Tal A. 2002. Pollution in a Promised Land. Berkeley (CA): University of California Press.
- ter Braak CJF and Smilauer P. 2006. CANOCO v4.54 Biometris. Wageningen, The Netherlands: Plant Research International.
- Theobald DM. 2004. Placing exurban land-use change in a human modification framework. Frontiers in Ecology and the Environment 2: 139–144.
- Trammel EJ, Weisberg PJ, and Bassett S. 2011. Avian response to urbanization in the arid riparian context of Reno, USA. Landscape and Urban Planning 102(2): 93–101.
- Woodroffe R and Ginsberg JR. 1998. Edge effects and the extinction of populations inside protected areas. Science 280(5372): 2126–2128.
- Yom Tov Y and Mendelsohn H. 1988. Changes in the distribution and abundance of vertebrates in Israel. In: Yom Tov Y and Tchernov E (Eds). The zoogeography of Israel: The distribution and abundance of a zoogeographical crossroad. Dordrecht: DRW Junk Publishers.



 מהדורה מודפסת
מהדורה מודפסת








